
Fertile island effect characteristics of two types of shrub-encroached grasslands in Xilingol, Inner Mongolia
Received date: 2025-04-25
Revised date: 2025-09-04
Online published: 2026-03-12
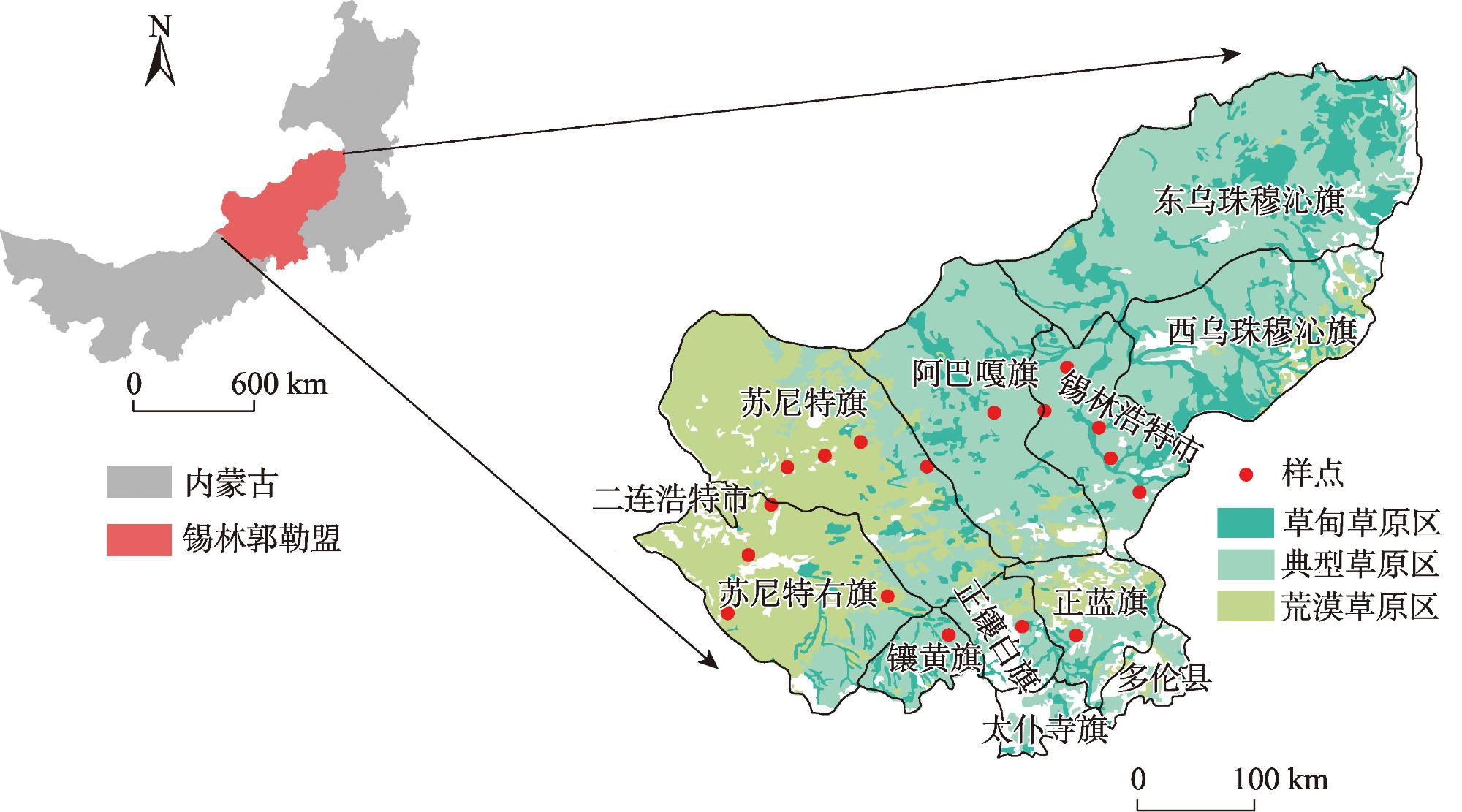
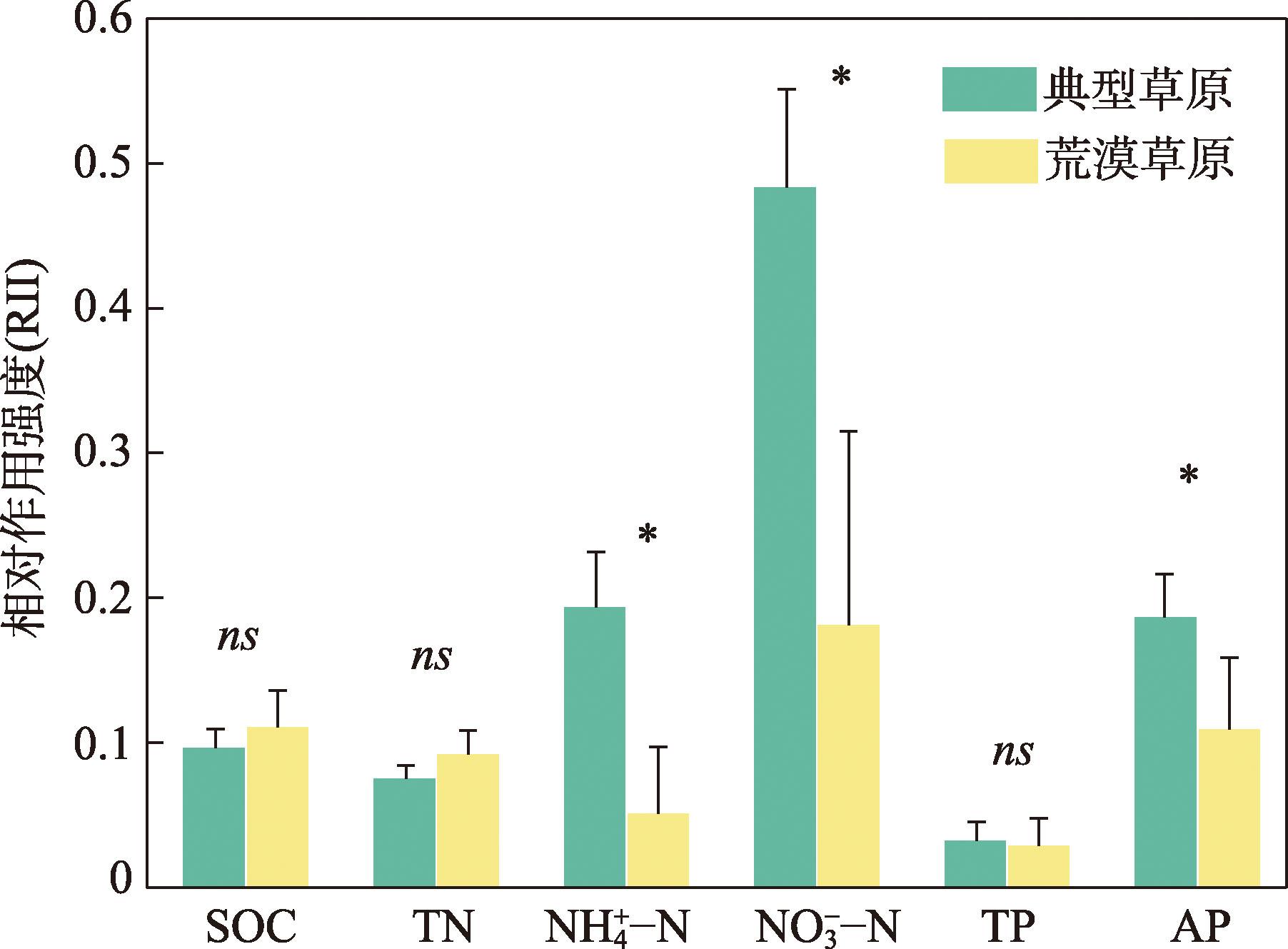
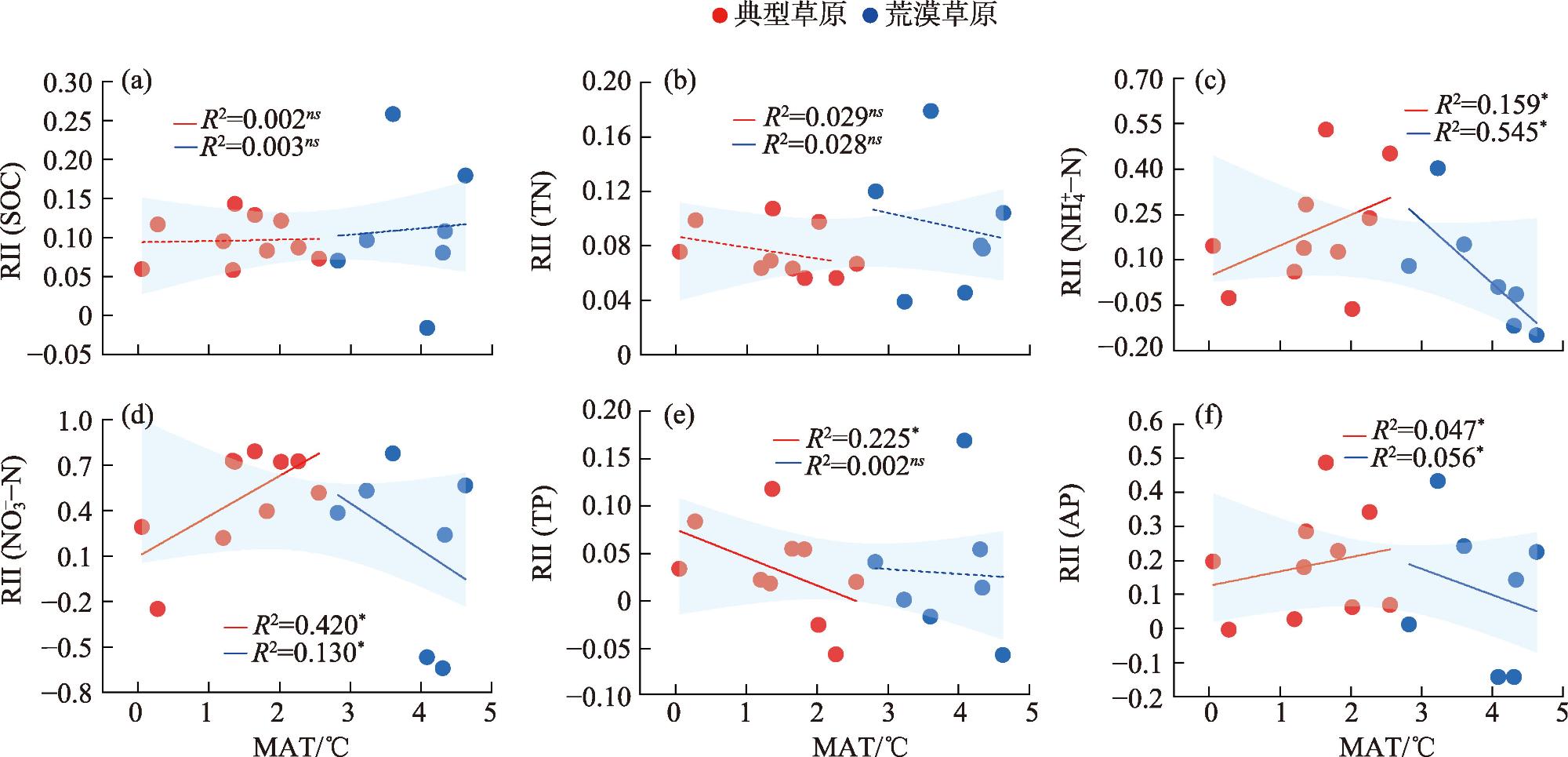
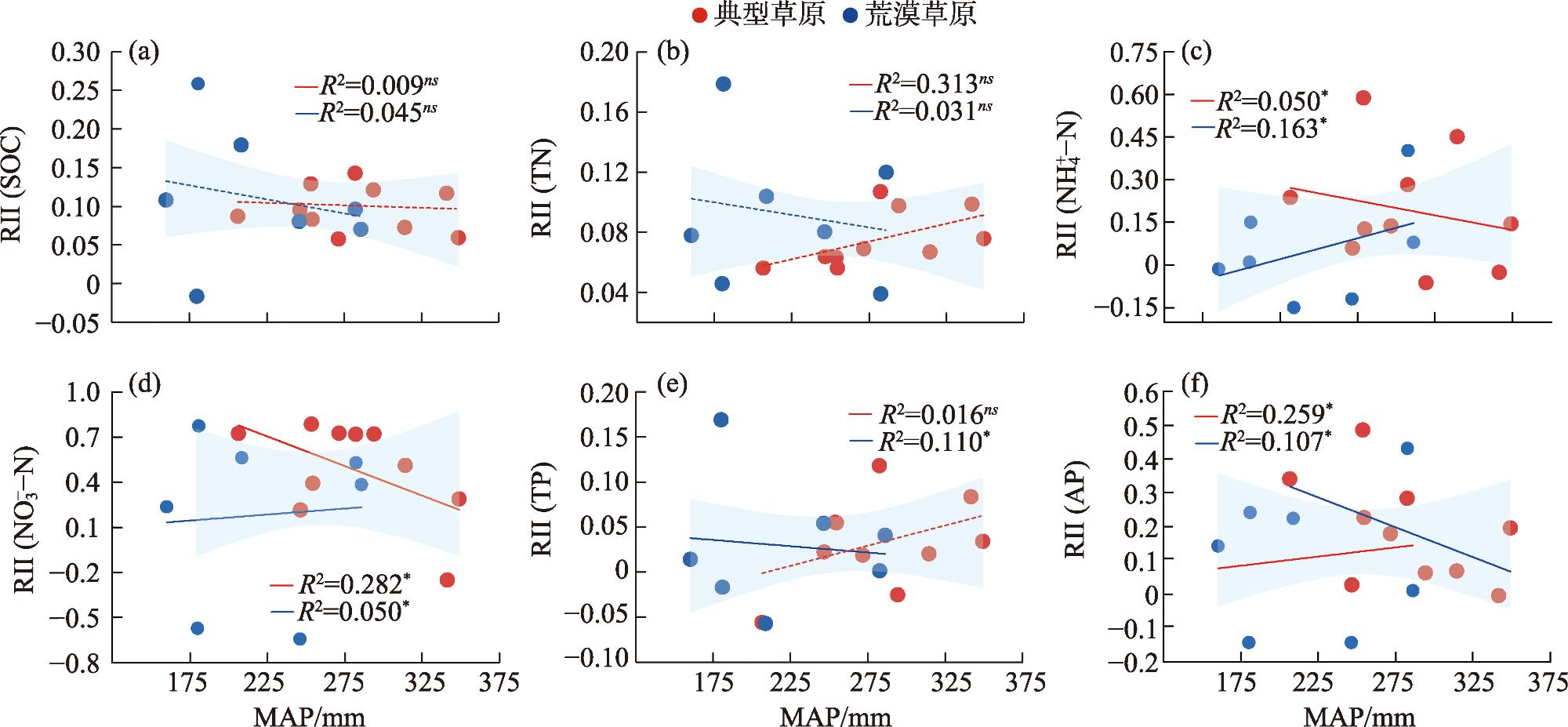
Shrub encroachment is widespread in arid and semi-arid regions, and it produces a fertility island effect, whereby soil nutrients under shrub patches become enriched, thereby influencing ecosystem nutrient cycling and structural stability. We selected 17 shrub-encroached grassland sites in typical grassland and desert-steppe regions of Xilingol League, Inner Mongolia and analyzed the soil stoichiometry of carbon (C), nitrogen (N) and phosphorus (P). By combining climatic data with structural equation modeling, we examined the characteristics of the fertility island effect under the two grassland types and the influence of climatic factors. Following shrub encroachment, C, N, and P concentrations under shrub patches were significantly higher than those in the herbaceous matrix (all P<0.05), indicating a pronounced fertility island effect. The relative interaction intensity (RII) of the fertility island effect was greater in the typical grassland region than in the desert-steppe region, with significant between-region differences in the RIIs of ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), and available P (P<0.05). The RIIs of different soil elements exhibited non-linear responses to mean annual temperature (MAT) and mean annual precipitation (MAP). In the typical grassland region, the RIIs of NH4+-N, NO3--N and available P increased significantly with MAT (all P<0.05), whereas in the desert-steppe region, these RIIs significantly decreased with increasing MAT (all P<0.05). The relationships of the RIIs of these three elements with MAP exhibited opposite patterns to those with temperature. Excluding the RII of available P, which was negatively correlated with aboveground biomass, the RIIs of the other elements were positively correlated with aboveground biomass. This study elucidated the fertility island effects induced by shrub encroachment and their relationships with climate and grassland type, providing a scientific basis for the ecological restoration of shrub-encroached grasslands and management of C, N, and P biogeochemical cycles.
FAN Shuxiang , GUO Jinyan , LI Huanhuan , LI He . Fertile island effect characteristics of two types of shrub-encroached grasslands in Xilingol, Inner Mongolia[J]. Arid Zone Research, 2025 , 42(12) : 2268 -2277 . DOI: 10.13866/j.azr.2025.12.10
表1 研究区基本信息Tab. 1 Basic information of the study area |
| 样地 设置 | 本底草原类型 | 年均温/℃ | 年降水量/mm | 地上生物量/(g·m-2) | 灌丛盖度/% |
|---|---|---|---|---|---|
| 样地1 | 典型草原 | 0.05 | 349.25 | 26.13 | 22 |
| 样地2 | 典型草原 | 0.28 | 341.60 | 31.13 | 16 |
| 样地3 | 典型草原 | 2.02 | 294.18 | 22.53 | 20 |
| 样地4 | 典型草原 | 1.65 | 253.78 | 34.86 | 21 |
| 样地5 | 典型草原 | 1.82 | 254.63 | 44.16 | 19 |
| 样地6 | 典型草原 | 1.37 | 282.49 | 45.01 | 14 |
| 样地7 | 典型草原 | 1.34 | 271.73 | 20.39 | 21 |
| 样地8 | 典型草原 | 1.21 | 246.67 | 16.04 | 20 |
| 样地9 | 典型草原 | 2.27 | 206.64 | 21.41 | 23 |
| 样地10 | 荒漠草原 | 3.60 | 180.90 | 47.51 | 18 |
| 样地11 | 荒漠草原 | 4.09 | 180.17 | 29.29 | 20 |
| 样地12 | 荒漠草原 | 4.63 | 208.70 | 21.25 | 12 |
| 样地13 | 荒漠草原 | 4.34 | 160.17 | 32.07 | 17 |
| 样地14 | 荒漠草原 | 4.31 | 246.38 | 38.97 | 12 |
| 样地15 | 荒漠草原 | 2.82 | 286.17 | 65.50 | 18 |
| 样地16 | 荒漠草原 | 3.23 | 282.62 | 21.11 | 11 |
| 样地17 | 典型草原 | 2.56 | 314.43 | 23.25 | 19 |
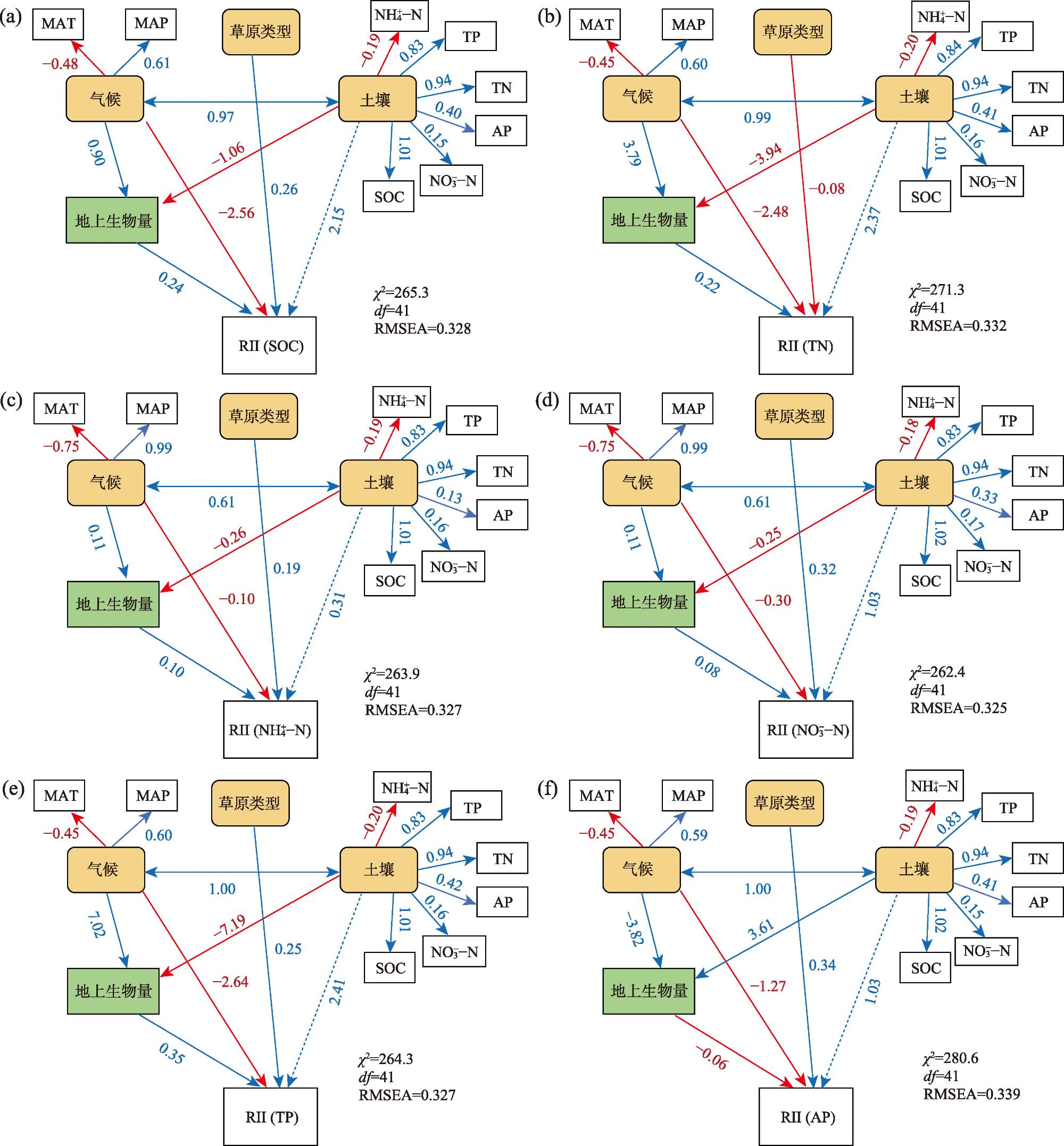
图6 沃岛效应气候、土壤、草原类型、地上生物量因子对各元素RII的结构方程模型注:直线箭头上的数字代表通径系数,系数越大相关性越强。蓝色直线箭头代表正相关,红色直线箭头代表负相关,实线代表相关性显著(P<0.05),虚线代表相关性不显著(P>0.05)。矩形框代表观测变量,圆角矩形代表潜变量。χ2为卡方值,df为自由度,RMSEA为均方根近似误差。 Fig. 6 Structural equation model of the effects of climate, soil, grassland type, and aboveground biomass factors on the RII of each element |
| [1] |
|
| [2] |
|
| [3] |
邢媛媛, 王永东, 雷加强. 草地灌丛化对植被与土壤的影响[J]. 干旱区研究, 2017, 34(5): 1157-1163.
[
|
| [4] |
熊小刚, 韩兴国. 内蒙古半干旱草原灌丛化过程中小叶锦鸡儿引起的土壤碳、氮资源空间异质性分布[J]. 生态学报, 2005, 25(7): 1678-1683.
[
|
| [5] |
|
| [6] |
|
| [7] |
白丽丽, 王文颖, 德却拉姆, 等. 祁连山典型植被土壤碳、氮、磷含量及生态化学计量特征的垂直变化[J]. 干旱区研究, 2024, 41(3): 444-455.
[
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
陈蕾伊, 沈海花, 方精云. 灌丛化草原: 一种新的植被景观[J]. 自然杂志, 2014, 36(6): 391-396.
[
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
徐浩然, 俞富洋, 贾聪慧, 等. 两种灌丛化草原小叶锦鸡儿的叶片化学计量特征[J]. 草地学报, 2021, 29(10): 2191-2199.
[
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
张敬敏, 珠娜, 蔡育蓉, 等. 小叶锦鸡儿(Caragana microphylla)灌丛对草本群落特征的影响[J]. 生态学报, 2023, 43(21): 8830-8839.
[
|
| [23] |
贾慎修. 中国草地区划的商讨[J]. 自然资源, 1985(2): 1-13.
[
|
| [24] |
|
| [25] |
|
| [26] |
何东健, 杨成海, 杨青, 等. 面向精准农业的农田土壤成分实时测定研究进展[J]. 农业工程学报, 2012, 28(7): 78-85.
[
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}