
Physiological, biochemical and morphological responses of Haloxylon ammodendron and Calligonum caput-medusae to drought stress
Received date: 2023-12-09
Revised date: 2024-04-18
Online published: 2025-08-14
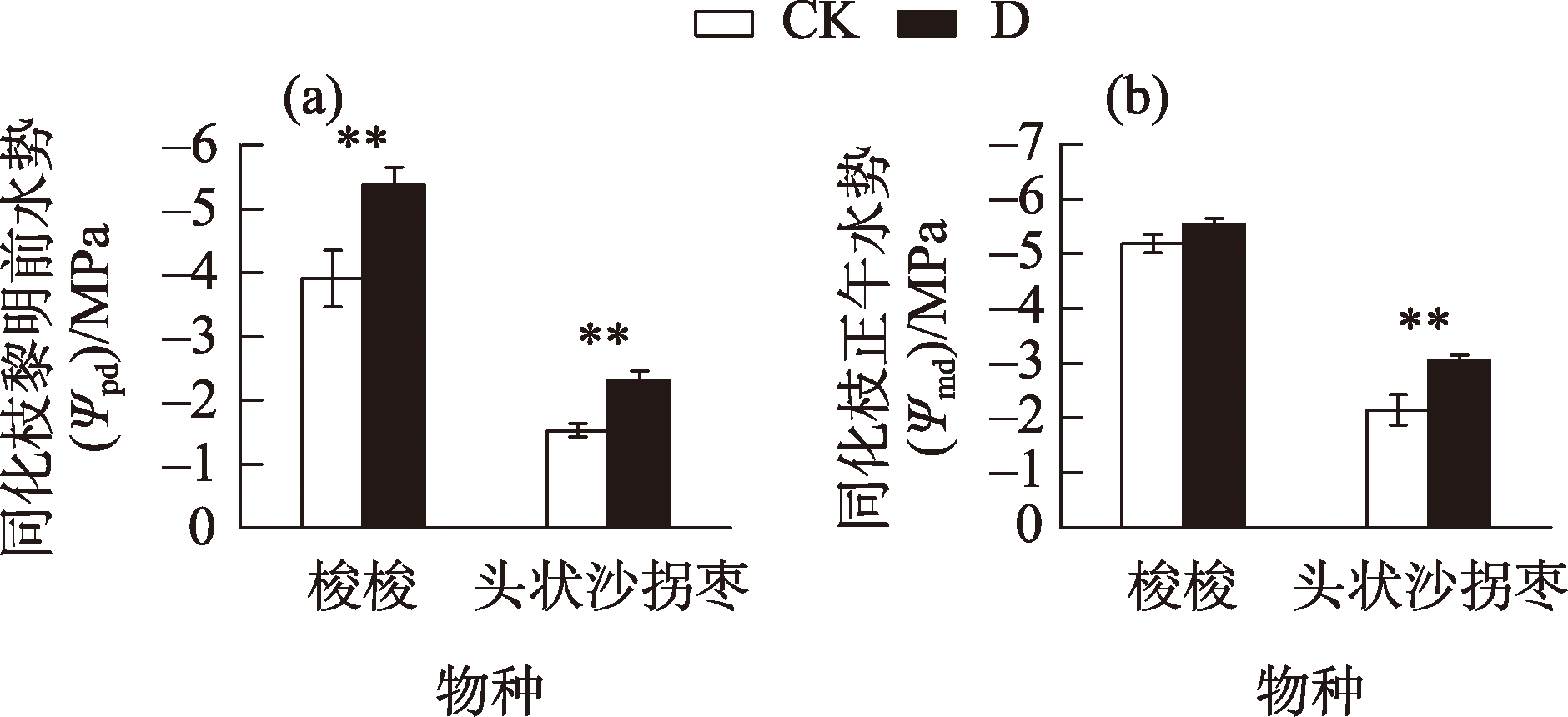
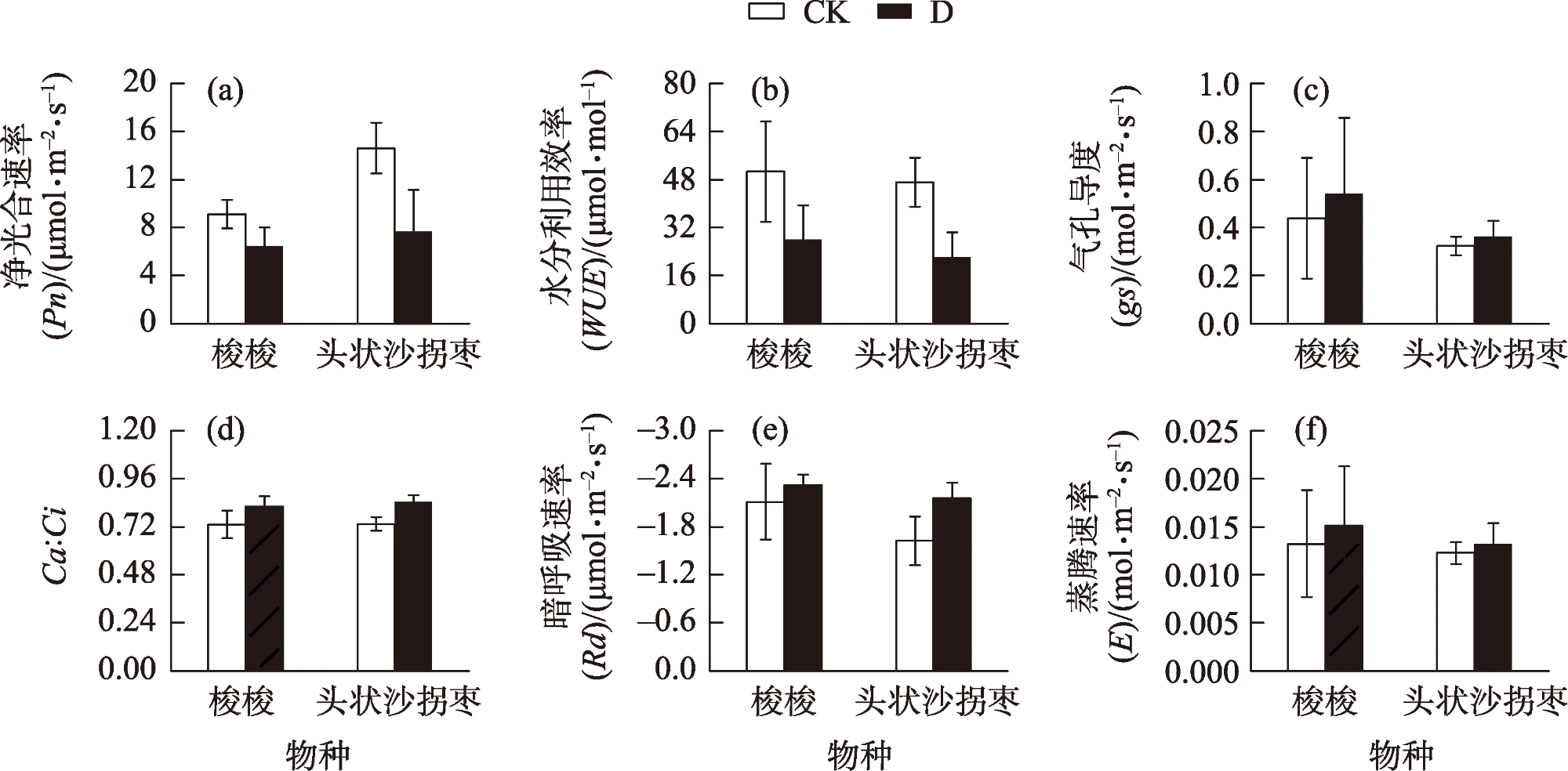
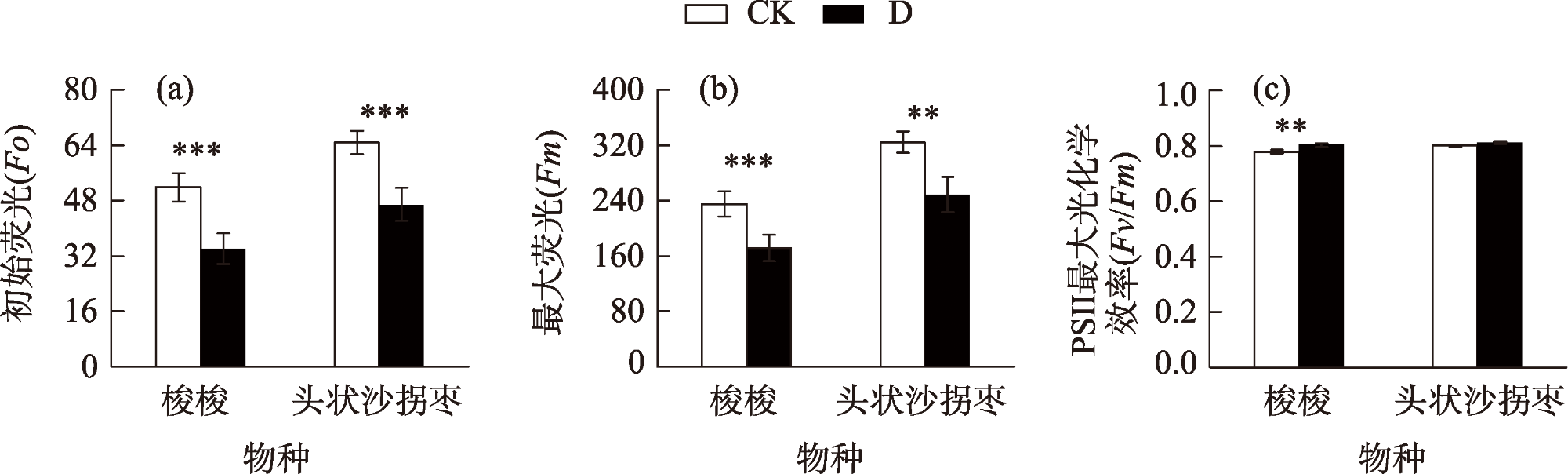
Desertification and drought have emerged as global ecological issues, which pose significant difficulties for plant adaptability and survival. Consequently, it is crucial to investigate the adaptation mechanisms of plants to drought stress to preserve and grow sustainably artificial ecosystems in desert regions. This study examined the physiological, biochemical, and morphological responses of Haloxylon ammodendron and Calligonum caput-medusae in the Taklimakan Desert shelterbelt by employing two treatments: irrigation (20-day irrigation cycle) (CK) and drought (D). The findings demonstrated that under drought stress, the assimilatory branches of both plants were noticeably shorter in terms of morphological traits. The branches of both were also significantly thinner. Regarding physiological reactions, there were no notable distinctions between the two in terms of the photosynthetic gas exchange parameters and the leaf dry mass during drought stress. Additionally, the carboxylation processes, metabolic reaction rates, chlorophyll a, chlorophyll b, total chlorophyll, and photosynthetic enzyme contents of both did not exhibit any appreciable alterations under drought stress. However, both showed substantially reduced Fm and Fo values. The Fv/Fm values of H. ammodendron were noticeably greater, but those of C. caput-medusae did not change much. According to our study, H. ammodendron and C. caput-medusae were able to adapt to drought by reducing the assimilate-branch growth, excitation capacity, and photosynthetic pigment activity, thereby preserving photosynthetic capacity. The photochemical system of C. caput-medusae was less vulnerable to damage due to a stable chlorophyll concentration.
ZHANG Bin , LI Congjuan , Yi Guangping , LIU Ran . Physiological, biochemical and morphological responses of Haloxylon ammodendron and Calligonum caput-medusae to drought stress[J]. Arid Zone Research, 2024 , 41(7) : 1177 -1184 . DOI: 10.13866/j.azr.2024.07.09
表1 不同干旱处理下同化枝形态及功能性状Tab. 1 Functional traits of assimilated branches under different drought treatments |
| 物种 | 干旱处理 | 土壤含水量 (SWC)/% | 同化枝直径 (LD)/mm | 同化枝长度 (LL)/cm | 同化枝相对含水量 (RWC)/% | 同化枝干物质量 (LDMC)/% |
|---|---|---|---|---|---|---|
| 梭梭 H. ammodendron | D | 0.04(0.01)* | 0.72(0.03)* | 1.57(0.12)** | 74(6.40) | 35(4.00) |
| CK | 0.71(0.41) | 0.81(0.03) | 2.84(0.15) | 82(1.30) | 54(1.00) | |
| 头状沙拐枣 C. caput-medusae | D | 0.08(0.02)* | 1.01(0.03)** | 5.11(0.48)** | 69(2.00)* | 39(1.60)* |
| CK | 0.63(0.12) | 1.39(0.03) | 10.01(0.37) | 78(0.60) | 32(2.00) |
注:*表示同一物种在不同干旱处理下存在显著性差异。*表示显著(P<0.05),**表示极显著(P<0.01),表中数据为均值(标准误),下同。 |
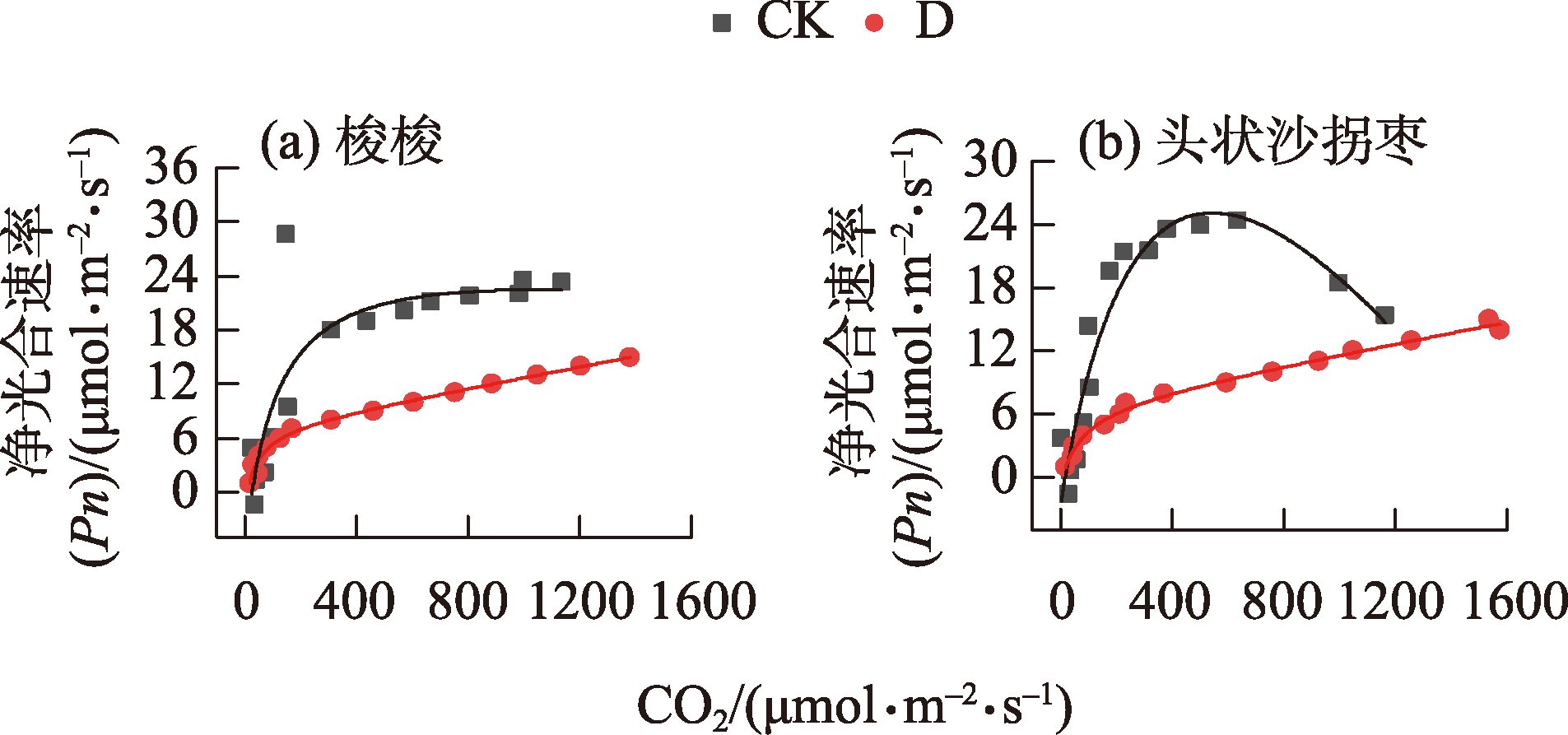
表2 不同干旱处理下A/Ca曲线参数Tab. 2 Parameters of A/Ca curves under different drought treatments |
| 物种 | 干旱处理 | 光合能力(Pnmax) /(μmol CO2·m-2·s-1) | 初始羧化效率(CE) /(mol·m-2·s-1) | CO2补偿点(CCP) /(μmol·m-1) | CO2饱和点(CSP) /(μmol·m-1) | 光呼吸速率(Rp) /(μmol CO2·m-2·s-1) |
|---|---|---|---|---|---|---|
| H. ammodendron | D | 14.53(2.85) | 0.11(0.05) | 59.33(14.79) | 566.33(106.67) | 4.89(1.39) |
| CK | 25.83(7.78) | 0.20(0.08) | 31.18(23.99) | 554.71(113.33) | 2.45(1.16) | |
| C. caput-medusae | D | 23.05(7.23) | 0.45(0.19) | 44.43(25.45) | 1038.61(435.63) | 13.39(9.00) |
| CK | 18.17(4.11) | 0.17(0.01) | 34.85(8.00) | 1236.23(427.50) | 4.66(0.77) |
图3 不同干旱处理下A/Ca曲线注:拟合同一物种不同干旱处理的A/Ca曲线,曲线均为三次测量的平均值。 Fig. 3 A/Ca curves under different drought treatments |
表3 不同干旱处理下同化枝光合酶及叶绿素含量Tab. 3 Photosynthetic chlorophyll traits of assimilated branches under different drought treatments |
| 物种 | 干旱处理 | 光合酶 (Rubisco)/(nmol·g-1·min-1) | 叶绿素a (Chla)/(mg·g-1) | 叶绿素b (Chlb)/(mg·g-1) | 总叶绿素 (Chl)/(mg·g-1) |
|---|---|---|---|---|---|
| H. ammodendron | D | 5.40(0.86) | 0.10(0.03) | 0.02(0.00) | 0.11(0.03) |
| CK | 7.11(0.76) | 0.15(0.02) | 0.01(0.00) | 0.17(0.03) | |
| C. caput-medusae | D | 269.00(42.43) | 0.24(0.02) | 0.05(0.01) | 0.29(0.21) |
| CK | 166.28(25.87) | 0.26(0.03) | 0.06(0.01) | 0.32(0.04) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
魏圆慧, 梁文召, 韩路, 等. 胡杨叶功能性状特征及其对地下水埋深的响应[J]. 生态学报, 2021, 41(13): 5368-5376.
[
|
| [6] |
徐梦琦, 高艳菊, 张志浩, 等. 干旱胁迫对疏叶骆驼刺幼苗生长和生理的影响[J]. 干旱区研究, 2023, 40(2): 257-267.
[
|
| [7] |
|
| [8] |
周洁, 杨晓东, 王雅芸, 等. 梭梭和骆驼刺对干旱的适应策略差异[J]. 植物生态学报, 2022, 46(9): 1064-1076.
[
|
| [9] |
|
| [10] |
|
| [11] |
何远政, 黄文达, 王怀海, 等. 沙质草地3种优势植物叶片光合生理对增温和降水减少的响应[J]. 西北植物学报, 2022, 42(4): 684-693.
[
|
| [12] |
|
| [13] |
|
| [14] |
李民青, 周乐, 王喜勇, 等. 7种荒漠木本植物枝干与叶片光合特征及其影响因素[J]. 应用生态学报, 2023, 34(10): 2637-2643.
[
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
闫海龙, 张希明, 许浩, 等. 塔里木沙漠公路防护林3种植物光合特性对干旱胁迫的响应[J]. 生态学报, 2010, 30(10): 2519-2528.
[
|
| [20] |
|
| [21] |
丁新原, 周智彬, 徐新文, 等. 咸水滴灌下塔克拉玛干沙漠腹地人工绿地土壤水分三维时空动态[J]. 应用生态学报, 2015, 26(9): 2600-2608.
[
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006: 77-74.
[
|
| [26] |
叶子飘, 于强. 光合作用对胞间和大气CO2响应曲线的比较[J]. 生态学杂志, 2009, 28(11): 2233-2238.
[
|
| [27] |
|
| [28] |
|
| [29] |
李炎, 王丹. 不同土壤水分测定方法的比较研究[J]. 安徽农业科学, 2010, 38(17): 9110-9112.
[
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
陈春晓, 谢秀华, 王宇鹏, 等. 盐分和干旱对沙枣幼苗生理特性的影响[J]. 生态学报, 2019, 39(12): 4540-4550.
[
|
| [34] |
闫海龙, 张希明, 许浩, 等. 塔里木沙漠公路防护林植物沙拐枣气体交换特性对干旱胁迫的响应[J]. 中国沙漠, 2007, 27(3): 460-465.
[
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
庞进平, 王永生. 油菜幼苗光合及叶绿素荧光参数对干旱胁迫的响应及其抗旱性分析[J]. 西北植物学报, 2023, 43(2): 276-284.
[
|
| [40] |
丁效东, 张士荣, 刘阳超, 等. 真盐生植物梭梭和囊果碱蓬幼苗耐干旱能力的研究[J]. 草业学报, 2015, 24(11): 240-246.
[
|
| [41] |
|
| [42] |
朱成刚, 陈亚宁, 李卫红, 等. 干旱胁迫对胡杨PSII光化学效率和激能耗散的影响[J]. 植物学报, 2011, 46(4): 413-424.
[
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}