
Effects of meteorological factors on the litter decomposition of the fungal endophyte Stipa purpurea symbiont in alpine grassland
Received date: 2025-03-27
Revised date: 2025-04-15
Online published: 2025-08-13
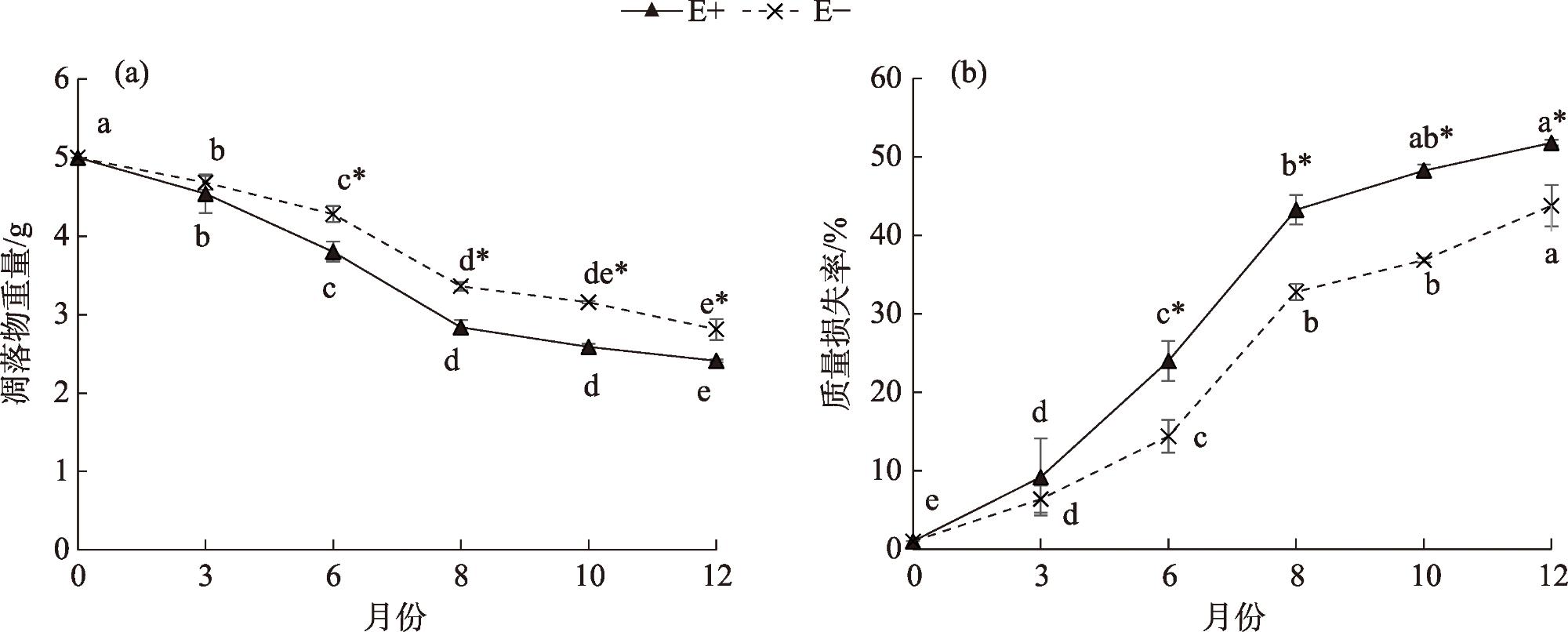
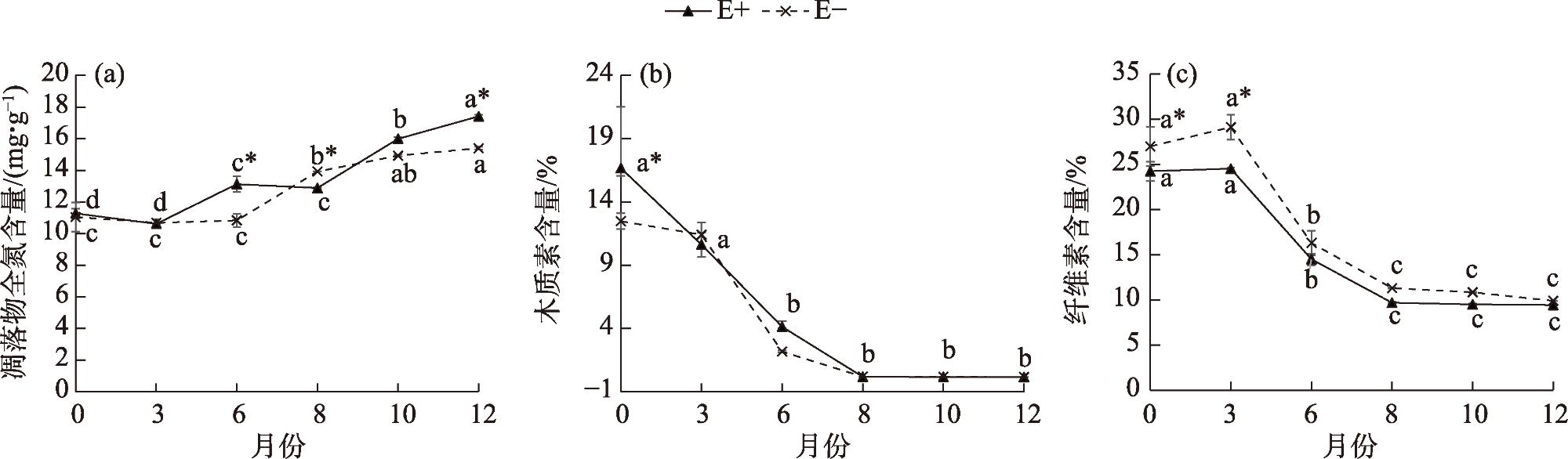
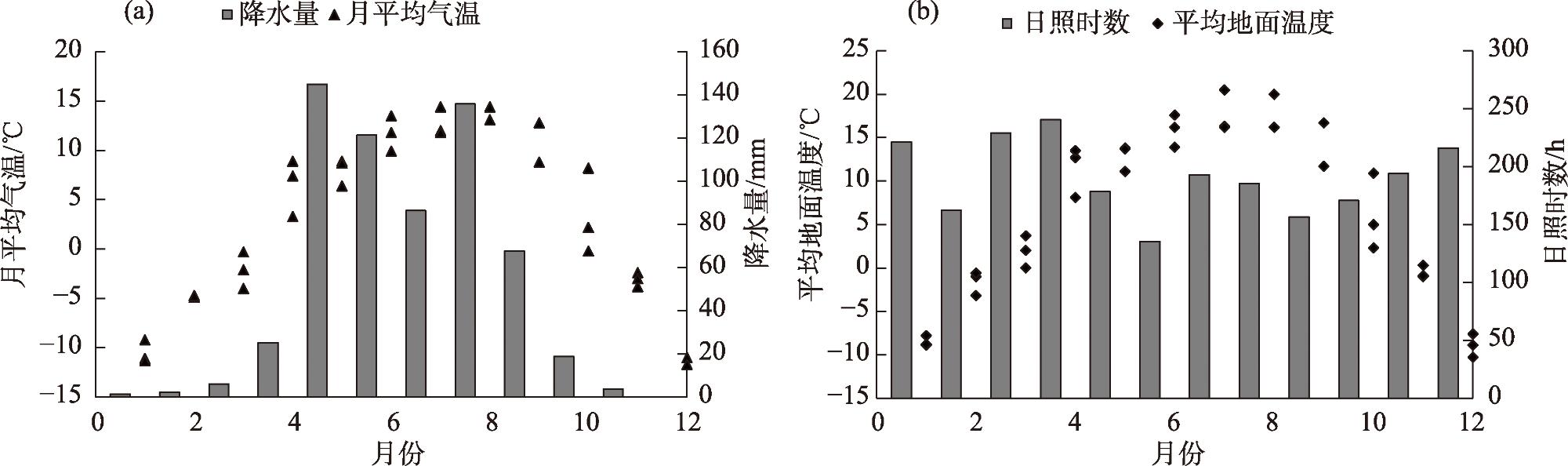
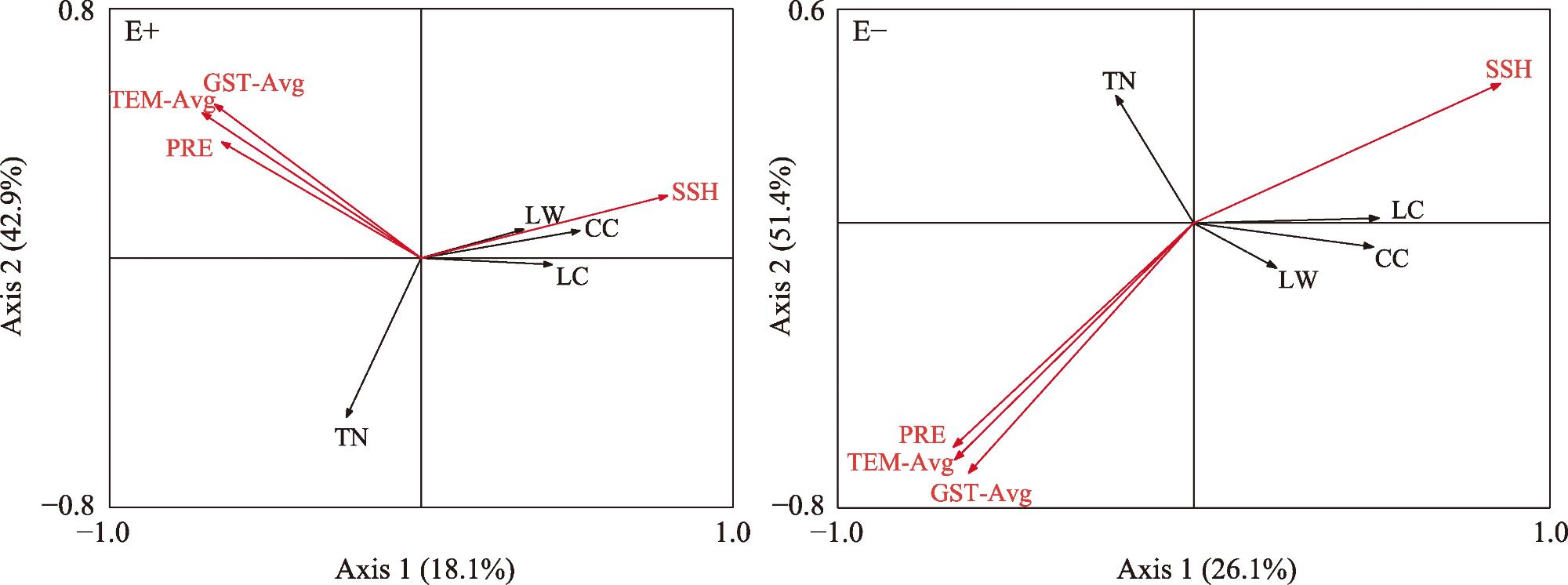
Litter decomposition has an important role in the carbon and nutrient cycling of terrestrial ecosystems. Climatic conditions are the main factors involved in litter decomposition. Currently, few studies have examined litter decomposition in alpine grassland ecosystems. To determine the effects of meteorological factors on the litter decomposition and nutrient release processes under a climate change background, the Stipa purpurea endophytic fungal symbiont was used along with the litter decomposition bag method to analyze the decomposition characteristics with endophytic fungi (E+) and without endophytic fungi (E-). The effect of meteorological factors on the decomposition rate of Stipa purpurea was analyzed. The results indicated that the decomposition rate of E+ was higher compared with that of E-, whereas the decomposition cycle was shorter. With the extension of time, the total nitrogen content of Stipa purpurea showed an increasing trend, lignin content gradually changed from significantly higher in E+ to no significant difference between the two, and the cellulose content gradually changed from significantly lower in E+ compared with E- (P<0.05). Regardless of endophytic fungi, the litter weight and mass loss rate of Stipa purpurea were correlated with the mean monthly temperature and the mean ground temperature (P<0.05). Precipitation was positively correlated with the litter decomposition rate of Stipa purpurea, and the total nitrogen content of the litter was positively correlated with temperature and precipitation (P<0.05). The lignin and cellulose content were negatively correlated with temperature and precipitation. The duration of sunshine had a positive effect on the decomposition of litter, and the lignin, cellulose, and litter weight content were strongly correlated with sunshine duration. Overall, endophytic fungi accelerate the decomposition of Stipa purpurea litter. For E+ and E-, the effect of meteorological factors on the decomposition of Stipa purpurea litter was consistent.
GUO Qiang , WANG Yuqin , SONG Meiling . Effects of meteorological factors on the litter decomposition of the fungal endophyte Stipa purpurea symbiont in alpine grassland[J]. Arid Zone Research, 2025 , 42(7) : 1269 -1278 . DOI: 10.13866/j.azr.2025.07.10
表1 E+和E-紫花针茅凋落物Olson指数模型和分解速率Tab. 1 Olson exponential model and decomposition rate of E+ and E- Stipa purpurea litters |
| 凋落物 种类 | 回归方程 | 相关系 数(R2) | 分解速 率(k) | ||
|---|---|---|---|---|---|
| E+ | y=1.1024e-0.869t | 0.970 | 0.869 | 0.80 | 3.45 |
| E- | y=1.09698e-0.66t | 0.963 | 0.66 | 1.05 | 4.54 |
注: 为凋落物分解50%所需要的时间; 为凋落物分解95%所需要的时间。 |
表2 紫花针茅凋落物分解特征与气象因子的相关性分析Tab. 2 Correlation analysis of litter decomposition characteristics and meteorological factors of Stipa purpurea |
| 凋落物种类 | 指标 | 月平均 气温 | 降水量 | 月平均地 面温度 | 日照时数 |
|---|---|---|---|---|---|
| E+ | 凋落物重量 | -0.990** | -0.802 | -0.993** | 0.290 |
| 质量损失率 | 0.989** | 0.805 | 0.992** | -0.287 | |
| 全氮 | 0.872* | 0.915* | 0.856* | -0.502 | |
| 木质素 | -0.938** | -0.659 | -0.955** | 0.169 | |
| 纤维素 | -0.924** | -0.703 | -0.935** | 0.065 | |
| E- | 凋落物重量 | -0.995** | -0.831* | -0.992** | 0.369 |
| 质量损失率 | 0.993** | 0.835* | 0.989** | -0.367 | |
| 全氮 | 0.931** | 0.885* | 0.917* | -0.398 | |
| 木质素 | -0.894* | -0.640 | -0.910* | 0.033 | |
| 纤维素 | -0.913* | -0.710 | -0.921** | 0.063 |
注:*表示P<0.05;**表示P<0.01。 |
| [1] |
王静, 赵萌莉,
[
|
| [2] |
王静, 张峰, 李治国, 等. 不同剂量凋落物添加对荒漠草原生物量的影响[J]. 中国草地学报, 2023, 45(6): 64-72.
[
|
| [3] |
曲浩, 赵学勇, 赵哈林, 等. 科尔沁沙地3种灌木凋落物分解速率及其与关键气象因子的关系[J]. 中国沙漠, 2010, 30(4): 844-849.
[
|
| [4] |
薛立, 邝立钢. 杉木凋落物分解速率的研究[J]. 四川林业科技, 1990, 11(1): 1-4.
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
李凯, 赵文东, 朱传晟, 等. 滨海防护林月凋落物量及其与气象因子的关系[J]. 森林与环境学报, 2021, 41(6): 570-575.
[
|
| [9] |
|
| [10] |
|
| [11] |
邓秀秀, 王忠诚, 李程, 等. 浙江天童常绿阔叶林凋落物量季节动态及其与气象因子的关系[J]. 中南林业科技大学学报, 2017, 37(3): 73-78.
[
|
| [12] |
宋梅玲, 王玉琴, 王宏生, 等. 内生真菌对高寒草地紫花针茅凋落物分解的影响[J]. 草业学报, 2021, 30(9): 150-158.
[
|
| [13] |
岳鹏鹏, 卢学峰, 叶润蓉, 等. 青海湖地区紫花针茅草原群落特征[J]. 草业学报, 2014, 23(4): 10-19.
[
|
| [14] |
张晓艳, 胡玉昆, 李凯辉, 等. 围封条件下紫花针茅群落主要结构特征和地上生物量变化[J]. 干旱区资源与环境, 2009, 23(1): 197-200.
[
|
| [15] |
洪江涛, 吴建波, 王小丹. 放牧和围封对藏北高寒草原紫花针茅群落生物量分配及碳、氮、磷储量的影响[J]. 草业科学, 2015, 32(11): 1878-1886.
[
|
| [16] |
付莉娇, 李雪琴, 范继辉, 等. 藏北高寒草原典型植物根际土壤细菌群落结构多样性及根系特征分析[J]. 草地学报, 2022, 30(5): 1131-1140.
[
|
| [17] |
鲍根生. 甘肃马先蒿寄生对紫花针茅内生真菌共生体生长和光合特性的影响[D]. 兰州: 兰州大学, 2015.
[
|
| [18] |
|
| [19] |
李春杰, 南志标, 刘勇, 等. 醉马草内生真菌检测方法的研究[J]. 中国食用菌, 2008, 27 (Suppl.): 16-19.
[
|
| [20] |
黎冬容, 张世庆, 甘世端, 等. 全自动凯氏定氮仪测定土壤全氮含量[J]. 南方国土资源, 2015(8): 38-39.
[
|
| [21] |
何忠武. 酸性洗涤纤维和木质素测定的国际标准[J]. 饲料广角, 2011(6): 31-32.
[
|
| [22] |
李娜, 赵传燕, 郝虎, 等. 海拔和郁闭度对祁连山青海云杉林叶凋落物分解的影响[J]. 生态学报, 2021, 41(11): 4493-4502.
[
|
| [23] |
|
| [24] |
王嘉年, 李向义, 李成道, 等. 自然光照和荫蔽条件下两种荒漠植物叶片凋落物分解特征研究[J]. 干旱区地理, 2023, 46(6): 949-957.
[
|
| [25] |
张悦, 张艺凡, 马怡波, 等. 森林生态系统凋落物分解影响因素研究进展[J]. 环境生态学, 2023, 5(4): 45-56.
[
|
| [26] |
|
| [27] |
|
| [28] |
侯卓男, 李欣彤, 张新军, 等. 海拔和坡向对色季拉山高山杜鹃凋落物分解的影响[J]. 中国农业大学学报, 2024, 29(4): 264-273.
[
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
宋学贵, 胡庭兴, 鲜骏仁, 等. 川西南常绿阔叶林凋落物分解及养分释放对模拟氮沉降的响应[J]. 应用生态学报, 2007, 18(10): 2167-2172.
[
|
| [34] |
|
| [35] |
|
| [36] |
彭少麟, 刘强. 森林凋落物动态及其对全球变暖的响应[J]. 生态学报, 2002, 22(9): 1534-1544.
[
|
| [37] |
|
| [38] |
周梦田, 刘莉, 付若仙, 等. 杉木与木荷凋落物分解对杉木人工林土壤碳氮含量和酶活性的影响[J]. 南京林业大学学报(自然科学版), 2024, 48(5): 131-138.
[
|
| [39] |
熊红福. 贵州省喀斯特地区凋落物分解对气候变暖的响应[J]. 现代农业科技, 2023(24): 118-121, 127.
[
|
| [40] |
|
| [41] |
代松家, 周晨霓, 段斐, 等. 组分和生境差异对藏东南原始冷杉林凋落物分解和养分释放特征的影响[J]. 中国水土保持科学, 2020, 18(6): 72-80.
[
|
| [42] |
霍利霞, 红梅, 赵巴音那木拉, 等. 氮沉降和降雨变化对荒漠草原凋落物分解的影响[J]. 生态学报, 2019, 39(6): 2139-2146.
[
|
| [43] |
|
| [44] |
王新源, 赵学勇, 李玉霖, 等. 环境因素对干旱半干旱区凋落物分解的影响研究进展[J]. 应用生态学报, 2013, 24(11): 3300-3310.
[
|
| [45] |
郑俊强, 郭瑞红, 李东升, 等. 氮沉降和干旱对阔叶红松林凋落物分解的影响[J]. 北京林业大学学报, 2016, 38(4): 21-28.
[
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}