
Effects of drought stress and rehydration on the physiological characteristics of Pinus sylvestris var. mongolica seedlings
Received date: 2024-05-26
Revised date: 2024-09-19
Online published: 2025-08-12
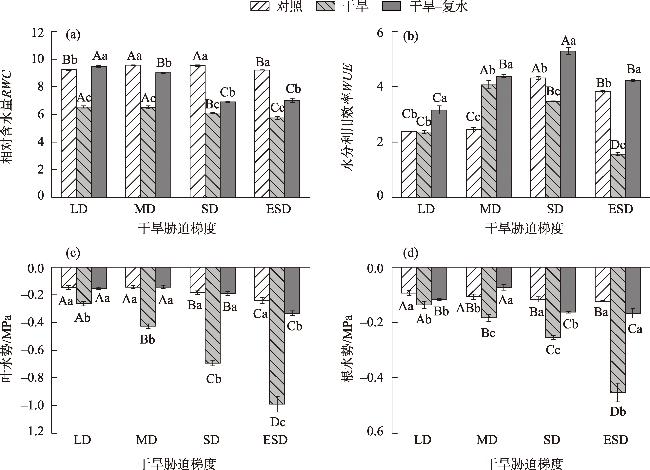
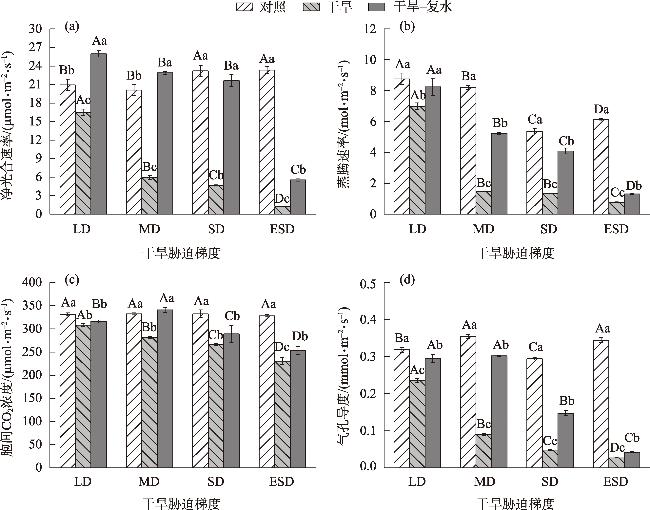
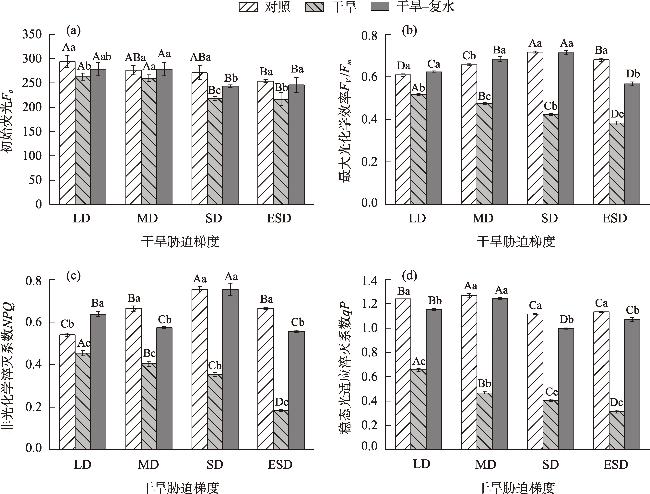
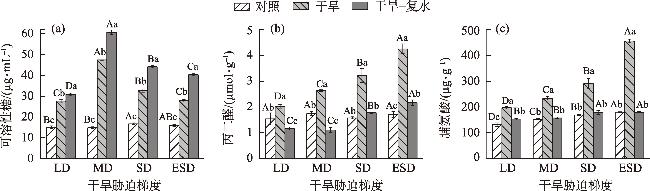
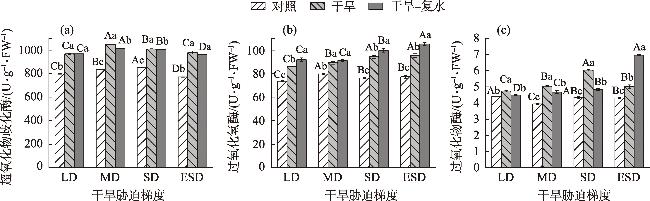
In this study, we investigated the physiological responses of Pinus sylvestris var. mongolica seedlings to drought stress and subsequent rehydration. A pot experiment was conducted using 2-year-old seedlings subjected to five water treatments, including control (80%), light drought (40%), moderate drought (20%), severe drought (10%), and extremely severe drought (5%). We measured leaf water content, photosynthesis, chlorophyll fluorescence, leaf osmoregulatory substances, and antioxidant enzyme activity during drought and after rehydration. The results showed the following: (1) The physiological state of the seedlings was minimally affected by the light drought. The activities of superoxide dismutase and peroxidase peaked under moderate and severe drought stress, increasing by 25.26% and 38.8%, respectively. Conversely, the net photosynthetic rate, transpiration rate, and photochemical quenching coefficient under extremely severe drought stress decreased by 94.76%, 87.19%, and 72.35%, respectively, while the leaf malondialdehyde content was the highest in this condition. (2) The chlorophyll fluorescence and leaf proline content of the seedlings were restored to control levels after rehydration. However, the average photosynthetic indices of rehydrated leaves only returned to 28.51% after extremely severe drought stress. Additionally, leaf water use efficiency, soluble sugar content, and antioxidant enzyme activity were significantly higher than the control after rehydration (P<0.05). In conclusion, extremely severe drought stress primarily reduces photosynthetic ability and disrupts the cell membrane stability of Pinus sylvestris var. mongolica seedlings. Conversely, rehydration after moderate drought improves water utilization and the scavenging ability of reactive oxygen species, thereby improving drought resistance. This improved information provides theoretical insights for the efficient cultivation, management, and evaluation of drought resistance in the Pinus sylvestris var. mongolica plantation.
WANG Zixiang , REN Yue , LU Ying , GAO Guanglei , DING Guodong , ZHANG Ying . Effects of drought stress and rehydration on the physiological characteristics of Pinus sylvestris var. mongolica seedlings[J]. Arid Zone Research, 2024 , 41(12) : 2120 -2131 . DOI: 10.13866/j.azr.2024.12.13
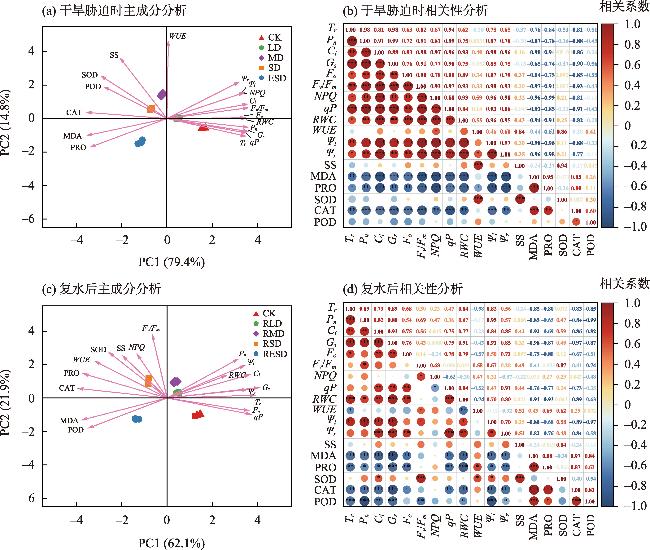
图6 干旱-复水条件下樟子松幼苗生理特征参数的主成分分析和相关性分析注:主成分分析中CK、RLD、RMD、RSD和RESD分别代表对照组、轻度干旱胁迫复水、中度干旱胁迫复水、重度干旱胁迫复水和极重度干旱胁迫复水;相关性分析中红色代表正相关关系,蓝色代表负相关关系;***、**、*分别表示Duncan法检验显著性小于0.001,0.01,0.05。 Fig. 6 Principal component analysis and correlation of physiological characteristics of Pinus sylvestris var. mongolica seedlings under drought stress conditions |
| [1] |
|
| [2] |
|
| [3] |
庞志强, 余迪求. 干旱胁迫下的植物根系-微生物互作体系及其应用[J]. 植物生理学报, 2020, 56(2): 109-126.
[
|
| [4] |
|
| [5] |
金思雨, 彭祚登. 刺槐和油松干旱胁迫响应研究进展[J]. 西北林学院学报, 2022, 37(4): 79-91.
[
|
| [6] |
|
| [7] |
赵娜, 赵旭, 李少宁, 等. 干旱复水下北京地区刺槐与油松叶绿素荧光特性研究[J]. 干旱地区农业研究, 2023, 41(2): 27-37.
[
|
| [8] |
曾露婧, 王国华. 干旱及复水对荒漠绿洲过渡带一年生草本植物生长及生理特性的影响[J]. 草业学报, 2024, 33(5): 41-57.
[
|
| [9] |
游韧, 邓湘雯, 胡彦婷, 等. 树木对干旱胁迫及复水的生理生态响应研究进展[J]. 林业科学, 2023, 59(11): 124-136.
[
|
| [10] |
路伟伟, 吴波, 白建华, 等. 樟子松人工林退化原因及研究展望[J]. 科学通报, 2023, 68(11): 1286-1297.
[
|
| [11] |
朱媛君, 山丹, 孙云海, 等. 天然樟子松林幼树更新格局及其影响因子分析[J]. 森林与环境学报, 2019, 39(4): 372-379.
[
|
| [12] |
|
| [13] |
|
| [14] |
王凯, 芦珊, 刘畅, 等. 土壤含水量对樟子松幼苗非结构性碳水化合物及生长的影响[J]. 生态学杂志, 2023, 42(3): 617-625.
[
|
| [15] |
宋立宁, 朱教君, 康宏樟. 樟子松幼苗水力结构参数和生长特征对模拟降水梯度的响应[J]. 干旱区研究, 2013, 30(6): 1021-1027.
[
|
| [16] |
|
| [17] |
何凌仙子, 贾志清, 刘涛, 等. 植物适应逆境胁迫研究进展[J]. 世界林业研究, 2018, 31(2): 13-18.
[
|
| [18] |
郭米山. 沙地樟子松外生菌根真菌特性及其对樟子松幼苗的影响[D]. 北京: 北京林业大学, 2020.
[
|
| [19] |
|
| [20] |
赵子涵, 王树森, 罗于洋, 等. 旱榆幼树对土壤失水及复水的生理响应过程研究[J]. 干旱区研究, 2022, 39(5): 1534-1542.
[
|
| [21] |
张玉玉, 王进鑫, 马戌, 等. 土壤干旱及复水对侧柏叶绿素荧光参数的影响[J]. 水土保持研究, 2021, 28(2): 242-247.
[
|
| [22] |
高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006: 1-231.
[
|
| [23] |
|
| [24] |
|
| [25] |
熊仕发, 吴立文, 陈益存, 等. 不同种源白栎幼苗叶片对干旱胁迫的响应及抗旱性评价[J]. 生态学杂志, 2020, 39(12): 3924-3933.
[
|
| [26] |
秦洁, 司建华, 贾冰, 等. 巴丹吉林沙漠典型植物水势与导水率的时空变化[J]. 生态学杂志, 2021, 40(6): 1629-1638.
[
|
| [27] |
陈斐, 闫霜, 王鹤龄, 等. 不同水分胁迫下的春小麦叶片气体交换参数和水分利用效率研究[J]. 干旱区研究, 2021, 38(3): 821-832.
[
|
| [28] |
魏媛, 喻理飞. 西南喀斯特地区构树苗木对土壤干旱胁迫的生理响应[J]. 水土保持研究, 2010, 17(2): 164-167.
[
|
| [29] |
周英, 吴沿友, 李海涛, 等. 土壤逐渐干旱条件下玉米和构树光合特性的变化[J]. 地球与环境, 2024, 52(3): 365-374.
[
|
| [30] |
|
| [31] |
付士磊, 周永斌, 何兴元, 等. 干旱胁迫对杨树光合生理指标的影响[J]. 应用生态学报, 2006, 17(11): 2016-2019.
[
|
| [32] |
|
| [33] |
冯延芝, 赵阳, 王保平, 等. 干旱复水对楸叶泡桐幼苗光合和叶绿素荧光的影响[J]. 中南林业科技大学学报, 2020, 40(4): 1-8.
[
|
| [34] |
高琦, 刘亚敏, 刘玉民, 等. 外源调节物质对干旱胁迫红椿苗木形态及光合生理的影响[J]. 西北农林科技大学学报(自然科学版), 2024, 52(7): 53-63.
[
|
| [35] |
杨肖华, 郭圣茂, 冯美玲, 等. 干旱胁迫及复水对射干光合作用和叶绿素荧光特性的影响[J]. 江西农业大学学报, 2018, 40(3): 525-532.
[
|
| [36] |
程爽,
[
|
| [37] |
冯蕊, 周琪, 吴令上, 等. PEG 6000模拟干旱胁迫对铁皮石斛幼苗生理和叶绿素荧光特性的影响[J]. 浙江农林大学学报, 2024, 41(1): 132-144.
[
|
| [38] |
李泽, 谭晓风, 卢锟, 等. 干旱胁迫对两种油桐幼苗生长、气体交换及叶绿素荧光参数的影响[J]. 生态学报, 2017, 37(5): 1515-1524.
[
|
| [39] |
刁珊,
[
|
| [40] |
李秋静, 卢广超, 薛立, 等. 臭氧与干旱胁迫对华南地区3种绿化树种荧光生理的影响[J]. 华南农业大学学报, 2015, 36(1): 91-95.
[
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
茶晓飞, 董琼, 段华超, 等. 干旱下白枪杆幼苗生物量及生理活性物质对钙添加的适应性调节[J]. 西北林学院学报, 2023, 38(3): 10-17.
[
|
| [45] |
张天泽, 孟凡君, 尹大川. 干旱胁迫下外生菌根菌对山新杨幼苗生物量、渗透调节物质和抗氧化酶的影响[J]. 菌物学报, 2023, 42(7): 1558-1574.
[
|
| [46] |
|
| [47] |
邓平, 吴敏, 林丁, 等. 干旱-复水对桂西北喀斯特地区青冈栎幼苗叶片光合能力、叶绿素荧光和显微结构的影响[J]. 西北植物学报, 2024, 44(1): 63-76.
[
|
| [48] |
|
| [49] |
|
| [50] |
赵英, 赵凯丽, 朱宇林, 等. 干旱胁迫与复水对喀斯特地区红背山麻杆生长及生理特性的影响[J]. 西北农林科技大学学报(自然科学版), 2023, 51(8): 59-68.
[
|
| [51] |
金思雨, 彭祚登. 刺槐苗木碳水生理参数对长期干旱及复水的响应变化[J]. 北京林业大学学报, 2023, 45(8): 43-56.
[
|
| [52] |
李春燕, 王进鑫, 薛设. 干旱及复水对侧柏幼苗叶片抗氧化酶活性的影响[J]. 西北林学院学报, 2015, 30(2): 33-37.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}