
Study on shrub species diversity and niche of wild fruit forest in Xinjiang
Received date: 2024-07-09
Revised date: 2024-08-30
Online published: 2025-08-12
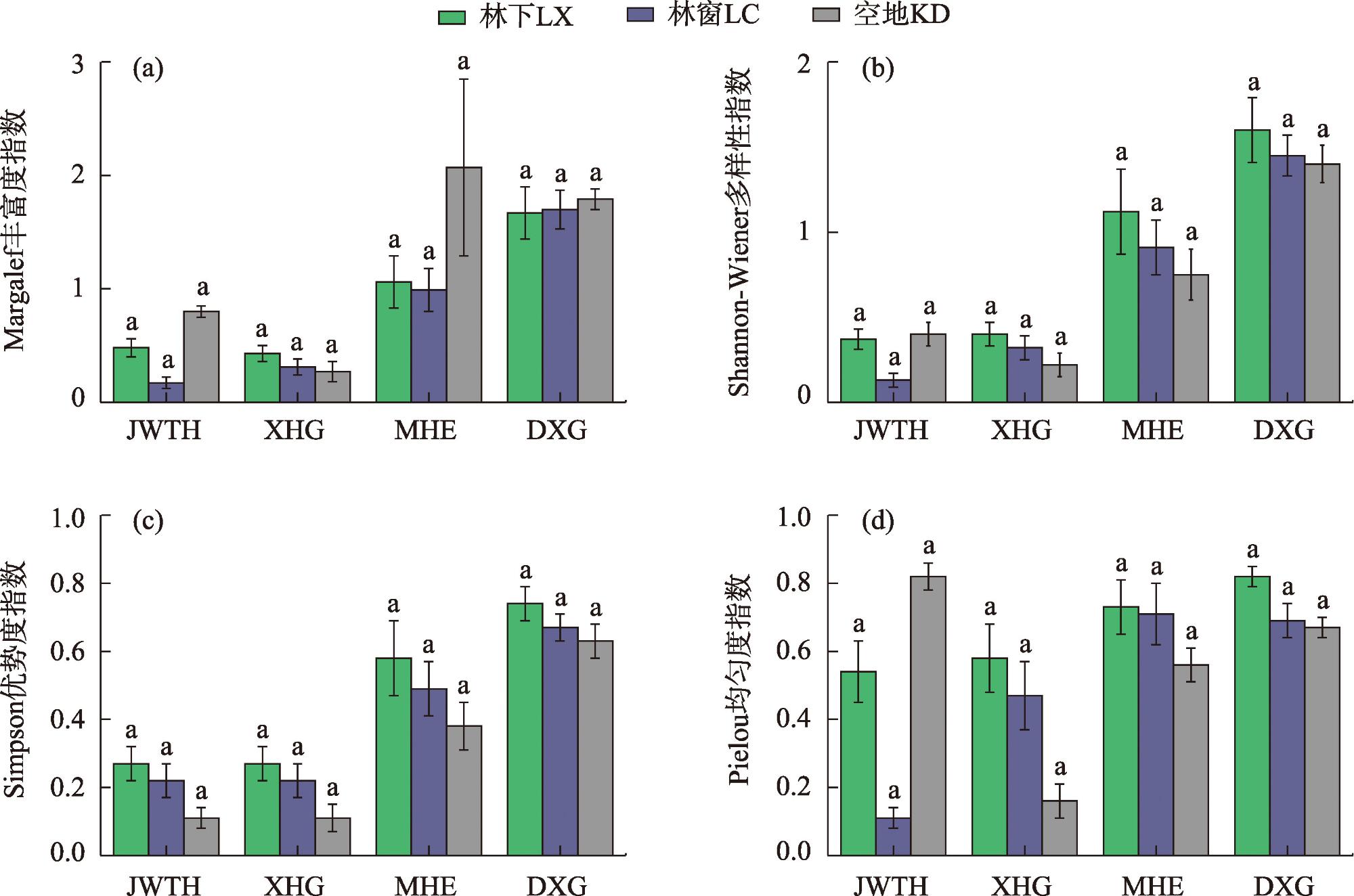
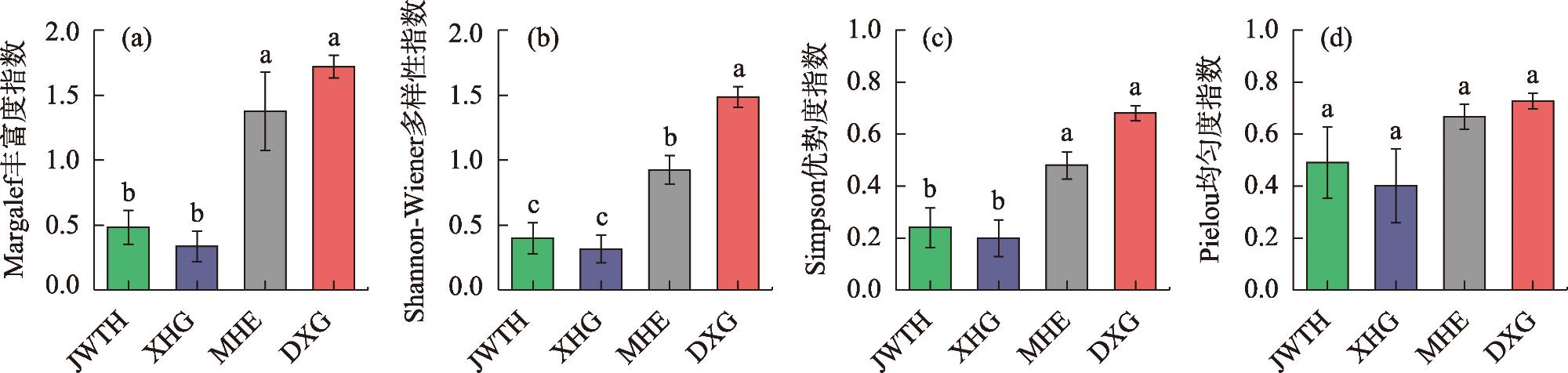
Shrub is an important part of forest community structure in Xinjiang wild fruit forest and plays an important role in maintaining ecosystem diversity and function. In this article, shrub communities of 4 wild fruit forest populations in JWTH, XHG, MHE and DXG were studied, then the species diversity and ecological niche of shrub were investigated by the means of sample plots (understory, forest gap, open space) and transversal lines (elevation lines between plots and wild fruit forest distribution). The results indicated that: (1) In the wild fruit forest: 110 species belonging to 26 genera and 18 families were found (see Annex 1 in the list), among which Rosaceae, Rosa and Cotoneaster are the dominant families and genera in the wild fruit forest; the areal-types of its families and genera are mainly distributed in the world and temperate zone; the important value accounts for a large proportion were Berberis (Berberis atrocarpa), Lonicera (Lonicera microphylla, Lonicera tatarica) and Rosa (Rosa beggeriana, Rosa laxa). There was no significant difference in shrub diversity coefficient among different habitats of the same wild fruit forest population, and the shrub diversity among populations gradually decreases from DXG, MHE, JWTH to XHG. (2) For the niche width, both BL and BS of B. atrocarpa in JWTH and MHE, L. microphylla in XHG were the largest, BL of B. atrocarpa in DXG had the largest Levins niche width and BS of Spiraea hypericifolia had the largest Shannon-Winner niche width. These species were widely distributed with strong ability of resource utilization and environmental adaptation. Ecological niche overlapped, <0.5 species pairs accounted for more than 50% of the total species pairs in wild fruit forest, and the similarity of environmental resources utilization was low, with weak competition between species. The shrubs of JWTH and MHE showed no significant negative association, while the shrubs of XHG and DXG showed no significant positive association, all of which were in the unstable early stage of succession. Conclusion: Xinjiang wild fruit forest was found with rich shrub species composition, the shrub species composition and diversity of different wild fruit forest populations were significantly different, the shrub population also was in an unstable state in the wild fruit forest ecosystem, which is closely related to the overall function of the wild fruit forest ecosystem.
Key words: shrub; diversity; flora; ecological niche; wild fruit forests; Xinjiang
WU Mingjiang , QIU Juan , ZHENG Feng , LING Xiaobo , WANG Xinyu , YANG Yang , YANG Jiaxin , LIU Liqiang . Study on shrub species diversity and niche of wild fruit forest in Xinjiang[J]. Arid Zone Research, 2024 , 41(12) : 2094 -2109 . DOI: 10.13866/j.azr.2024.12.11
表1 样地信息概况Tab. 1 Overview of plot information |
| 野果林居群 | 生境 | 编号 | 经度 | 纬度 | 海拔/m | 坡度/(°) | 坡向 |
|---|---|---|---|---|---|---|---|
| 交吾托海JWTH | 林下LX | LX-1 | 83°36′25.71″E | 43°22′35.49″N | 1473.7 | 26 | 西北322° |
| LX-2 | 83°33′56.12″E | 43°22′24.41″N | 1440.5 | 20 | 西北322° | ||
| LX-3 | 83°33′48.59″E | 43°22′17.69″N | 1459.8 | 17 | 西北318° | ||
| 林窗LC | LC-1 | 83°36′21.73″E | 43°22′37.14″N | 1446.7 | 20 | 北34° | |
| LC-2 | 83°36′25.22″E | 43°22′33.83″N | 1490.1 | 22 | 北20° | ||
| LC-3 | 83°35′50.52″E | 43°22′40.52″N | 1395.2 | 15 | 北13° | ||
| 空地KD | KD-1 | 83°36′20.90″E | 43°22′35.92″N | 1446.0 | 24 | 北345° | |
| KD-2 | 83°36′03.07″E | 43°22′37.76″N | 1420.7 | 16 | 北355° | ||
| KD-3 | 83°34′58.76″E | 43°22′36.76″N | 1392.1 | 15 | 东北25° | ||
| 杏花沟XHG | 林下LX | LX-1 | 83°26′14.58″E | 43°32′24.99″N | 1138.0 | 31 | 东北54° |
| LX-2 | 83°26′04.13″E | 43°32′56.92″N | 1326.5 | 29 | 东南154° | ||
| LX-3 | 83°26′18.58″E | 43°23′45.05″N | 1243.1 | 20 | 西南213° | ||
| 林窗LC | LC-1 | 83°26′04.30″E | 43°32′59.22″N | 1352.0 | 35 | 东107° | |
| LC-2 | 83°26′19.21″E | 43°32′36.39″N | 1138.5 | 8 | 东108° | ||
| LC-3 | 83°25′57.22″E | 43°33′13.23″N | 1568.5 | 42 | 东74° | ||
| 空地KD | KD-1 | 83°26′22.25″E | 43°32′50.53″N | 1291.5 | 22 | 南154° | |
| KD-2 | 83°26′21.93″E | 43°32′54.09″N | 1253.2 | 34 | 北307° | ||
| KD-3 | 83°25′56.84″E | 43°33′10.84″N | 1535.0 | 37 | 东南138° | ||
| 莫乎尔MHE | 林下LX | LX-1 | 82°43′13.77″E | 43°12′09.82″N | 1369.7 | 15 | 东81° |
| LX-2 | 82°43′33.43″E | 43°12′08.87″N | 1317.1 | 28 | 西北296° | ||
| LX-3 | 82°44′19.49″E | 43°14′03.57″N | 1261.0 | 23 | 北22° | ||
| 林窗LC | LC-1 | 82°43′33.19″E | 43°12′08.28″N | 1315.1 | 26 | 西南243° | |
| LC-2 | 82°44′09.85″E | 43°14′02.99″N | 1322.5 | 15 | 西南234° | ||
| LC-3 | 82°45′16.93″E | 43°13′20.79″N | 1242.2 | 25 | 东北57° | ||
| 空地KD | KD-1 | 82°43′13.33″E | 43°12′29.28″N | 1325.4 | 21 | 东83° | |
| KD-2 | 82°43′39.12″E | 43°14′15.33″N | 1330.2 | 16 | 东北25° | ||
| KD-3 | 82°45′25.44″E | 43°13′87.20″N | 1306.2 | 24 | 东北36° | ||
| 大西沟DXG | 林下LX | LX-1 | 80°46′49.10″E | 44°26′12.75″N | 1230.2 | 19 | 南182° |
| LX-2 | 82°45′17.19″E | 44°25′50.41″N | 1160.1 | 26 | 南153° | ||
| LX-3 | 80°46′19.63″E | 44.26°39.90′″N | 1253.9 | 27 | 西南205° | ||
| 林窗LC | LC-1 | 80°46′99.66″E | 44°26′13.06″N | 1243.1 | 21 | 南180° | |
| LC-2 | 80°46′38.51″E | 44°25′56.95″N | 1171.3 | 22 | 南183° | ||
| LC-3 | 80°46′42.35″E | 44°25′55.13″N | 1166.9 | 22 | 西南211° | ||
| 空地KD | KD-1 | 80°46′38.19″E | 44°25′51.66″N | 1187.6 | 29 | 东102° | |
| KD-2 | 80°46′36.23″E | 44°25′49.46″N | 1202.2 | 34 | 东96° | ||
| KD-3 | 80°47′35.33″E | 44°26′01.10″N | 1246.0 | 24 | 东南119° |
表2 新疆野果林灌木科、属的组成Tab. 2 The composition of shrub families and genera in Xinjiang wild fruit forest |
| 所含属数 | 科数 | 百分比 | 所含种数 | 属数 | 百分比/% |
|---|---|---|---|---|---|
| 含有1属的科 | 15 | 83.33 | 含有1~3种的属 | 16 | 61.54 |
| 含有2属的科 | 2 | 11.11 | 含有4~6种的属 | 4 | 15.38 |
| 含有3属及以上的科 | 1 | 5.56 | 含7种及以上的属 | 6 | 23.08 |
| 合计 | 18 | 100.00 | 合计 | 26 | 100.00 |
表3 野果林灌木植物科的分布区类型Tab. 3 The areal-types of shrub family in wild fruit forest |
| 区系组 | 类型及变型 | 科数 | 占比/% |
|---|---|---|---|
| 世界分布 | 1. 世界广布 | 8科(毛茛科Ranunculaceae、虎耳草科Saxifragaceae、蔷薇科Rosaceae、豆科Leguminosae、鼠李科Rhamnaceae、白花丹科Plumbaginaceae、蓼科Polygonaceae、茄科Solanaceae) | 44.44 |
| 热带成分 | 2. 泛热带(热带广布) | 2科(卫矛科Celastraceae、蒺藜科Zygophyllaceae) | 11.11 |
| (6d)南非(主要是好望角) | 1科(杜鹃花科Ericaceae) | 5.56 | |
| 温带成分 | 8. 北温带广布 | 1科(忍冬科Caprifoliaceae) | 5.56 |
| 8-4. 北温带和南温带间断分布 | 3科(杨柳科Salicaceae、槭树科Aceraceae、胡颓子科Elaeagnaceae) | 16.66 | |
| 8-5. 欧亚和南美洲温带间断 | 2科(麻黄科Ephedraceae、小檗科Berberidaceae) | 11.11 | |
| 10. 旧世界温带 | 1科(柽柳科Tamaricaceae) | 5.56 | |
| 合计 | - | 18科 | 100.00 |
表4 野果林灌木植物属的分布区类型Tab. 4 The areal-types of shrub genera in wild fruit forest |
| 区系组 | 类型及变型 | 属数 | 占比/% |
|---|---|---|---|
| 世界分布 | 1. 世界分布 | 3属(铁线莲属Clematis、悬钩子属Rubus、鼠李属Rhamnus) | 11.54 |
| 温带成分 | 8. 北温带分布 | 6属(山楂属Crataegus、蔷薇属Rosa、花楸属Sorbus、绣线菊属Spiraea、忍冬属Lonicera、荚蒾属Viburnum) | 23.08 |
| 8-2. 北极-高山 | 1属(北极果属Arctous) | 3.85 | |
| 8-4. 北温带和南温带间断(泛温带) | 7属(李属Prunus、柳属Salix、茶藨子属Ribes、卫矛属Euonymus、槭属Acer、萹蓄属Polygonum、枸杞属Lycium) | 26.90 | |
| 8-5. 欧亚和南温带间断 | 2属(小檗属Berberis、麻黄属Ephedra) | 7.69 | |
| 10. 旧世界温带分布 | 3属(栒子属Cotoneaster、水柏枝属Myricaria、沙棘属Hippophae) | 11.54 | |
| 10-1. 地中海、西亚(或中亚)和东亚间断 | 1属(木蓼属Atraphaxis) | 3.85 | |
| 11. 温带亚洲分布 | 1属(锦鸡儿属Caragana) | 3.85 | |
| 12-3. 地中海至温带-亚热带、大洋洲和南美洲间断 | 1属(白刺属Nitraria) | 3.85 | |
| 13. 中亚分布 | 1属(伊犁花属Ikonnikovia) | 3.85 | |
| 合计 | - | 26属 | 100.00 |
表5 野果林各居群灌木物种重要值Tab. 5 Important value of shrub species in each population of wild fruit forest |
| 物种 | 交吾托海JWTH | 杏花沟XHG | 莫乎尔MHE | 大西沟DXG | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 林下LX | 林窗LC | 空地KD | 居群 | 林下LX | 林窗LC | 空地KD | 居群 | 林下LX | 林窗LC | 空地KD | 居群 | 林下LX | 林窗LC | 空地KD | 居群 | ||||
| 黑果小檗Berberis atrocarpa | 11.28 | 6.00 | 11.32 | 28.60 | - | - | - | - | 14.05 | 16.73 | 10.19 | 40.97 | 9.66 | 7.29 | 4.54 | 21.49 | |||
| 小叶忍冬Lonicera microphylla | 5.87 | 10.56 | 3.53 | 19.96 | 8.93 | 3.23 | 10.71 | 22.88 | - | - | 2.03 | 2.03 | 1.73 | 0.66 | 1.85 | 4.24 | |||
| 矮小忍冬Lonicera humilis | - | 18.65 | 3.49 | 22.14 | 4.71 | 2.97 | 3.51 | 11.19 | 1.25 | - | 1.79 | 3.04 | - | - | - | - | |||
| 新疆忍冬Lonicera tatarica | - | - | - | - | - | - | 5.79 | 5.79 | 1.89 | 1.23 | 12.32 | 15.44 | 2.57 | - | 1.41 | 3.98 | |||
| 弯刺蔷薇Rosa beggeriana | - | 2.59 | 16.70 | 20.23 | - | - | - | - | 4.90 | 4.70 | 3.53 | 13.13 | 0.91 | 1.20 | - | 2.11 | |||
| 疏花蔷薇Rosa laxa | - | - | 3.54 | 2.59 | 3.42 | - | - | 3.42 | 5.10 | 2.64 | - | 7.74 | 2.06 | 2.36 | 1.43 | 5.85 | |||
| 密刺蔷薇Rosa spinosissima | - | - | - | - | - | - | - | - | - | - | 1.05 | 1.05 | - | - | 1.92 | 1.92 | |||
| 宽刺蔷薇Rosa platyacantha | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.66 | - | 0.66 | |||
| 腺毛蔷薇Rosa fedtschenkoana | - | - | - | - | - | - | - | - | 1.37 | - | - | 1.37 | - | - | - | - | |||
| 樱桃李Prunus cerasifera | 6.48 | - | - | 6.48 | - | - | - | - | - | - | - | - | - | 1.19 | 2.75 | 3.94 | |||
| 准噶尔栒子Cotoneaster soongoricus | - | - | - | - | - | - | - | - | 1.81 | 1.74 | 1.09 | 4.62 | 1.30 | 1.13 | 0.94 | 3.37 | |||
| 水栒子Cotoneaster multiflorus | - | - | - | - | - | - | - | - | 1.58 | - | - | 1.58 | 0.70 | - | 0.67 | 1.37 | |||
| 药鼠李Rhamnus cathartica | - | - | - | - | - | - | - | - | - | - | - | - | 3.13 | 2.26 | - | 5.39 | |||
| 天山樱桃Prunus tianshanica | - | - | - | - | - | - | - | - | - | - | - | - | - | 1.21 | 5.41 | 6.62 | |||
| 金丝桃叶绣线菊Spiraea hypericifolia | - | - | - | - | - | - | - | - | - | 1.42 | 1.07 | 2.50 | 3.65 | 8.57 | 17.65 | 29.88 | |||
| 黄刺条Caragana frutex | - | - | - | - | - | - | - | - | - | - | - | - | 2.23 | 2.77 | 4.18 | 9.18 | |||
| 沙棘Hippophae rhamnoides | - | - | - | - | - | 26.52 | 26.52 | 53.04 | - | - | - | - | - | - | - | - | |||
| 欧洲木莓Rubus caesius | - | - | - | - | - | 3.68 | - | 3.68 | - | - | - | - | - | - | - | - | |||
| 铁线莲Clematis florida | - | - | - | - | - | - | - | - | 4.01 | - | 2.52 | 6.53 | - | - | - | - | |||
| 和值 | 23.63 | 37.80 | 38.57 | 100.00 | 17.06 | 36.40 | 46.54 | 100.00 | 35.95 | 28.46 | 35.59 | 100.00 | 27.95 | 29.30 | 42.75 | 100.00 | |||
表6 4个野果林居群灌木生态位宽度Tab. 6 Shrub niche breadth of four wild fruit forest communities |
| 野果林居群 | 交吾托海JWTH | 杏花沟XHG | 莫乎尔MHE | 大西沟DXG | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 黑果小檗Berberis atrocarpa | 4.57 | 1.63 | - | - | 5.32 | 1.83 | 6.81 | 2.01 | |||
| 小叶忍冬Lonicera microphylla | 2.78 | 1.06 | 3.10 | 1.24 | 1.00 | 0.00 | 4.17 | 1.59 | |||
| 矮小忍冬Lonicera humilis | 1.09 | 0.18 | 2.33 | 0.96 | 1.80 | 0.64 | - | - | |||
| 新疆忍冬Lonicera tatarica | - | - | 1.00 | 0.00 | 1.40 | 0.54 | 3.85 | 1.47 | |||
| 弯刺蔷薇Rosa beggeriana | 2.45 | 0.99 | 2.47 | 1.24 | 5.05 | 1.69 | |||||
| 疏花蔷薇Rosa laxa | 1.00 | 0.00 | 1.00 | 0.00 | 2.35 | 1.01 | 5.05 | 1.69 | |||
| 密刺蔷薇Rosa spinosissima | - | - | - | - | 1.00 | 0.00 | 1.00 | 0.00 | |||
| 宽刺蔷薇Rosa platyacantha | - | - | - | - | - | - | 1.00 | 0.00 | |||
| 腺毛蔷薇Rosa fedtschenkoana | - | - | - | - | 1.00 | 0.00 | - | - | |||
| 樱桃李Prunus cerasifera | 1.80 | 0.64 | - | - | - | - | 4.8 | 1.59 | |||
| 准噶尔栒子Cotoneaster soongoricus | - | - | - | - | 2.33 | 0.96 | 4.26 | 1.52 | |||
| 水栒子Cotoneaster multiflorus | - | - | - | - | 1.00 | 0.00 | 1.6 | 0.56 | |||
| 药鼠李Rhamnus cathartica | - | - | - | - | - | - | 3.05 | 1.28 | |||
| 天山樱桃Prunus tianshanica | - | - | - | - | - | - | 2.84 | 1.29 | |||
| 金丝桃叶绣线菊Spiraea hypericifolia | - | - | - | - | 1.60 | 0.56 | 6.79 | 2.03 | |||
| 黄刺条Caragana frutex | - | - | - | - | - | - | 3.91 | 1.76 | |||
| 沙棘Hippophae rhamnoides | - | - | 2.00 | 0.69 | - | - | - | - | |||
| 欧洲木莓Rubus caesius | - | - | 1.00 | 0.00 | - | - | - | - | |||
| 铁线莲Clematis florida | - | - | - | - | 4.45 | 1.55 | - | - | |||
| 平均 | 2.28 | 0.75 | 1.74 | 0.48 | 2.14 | 0.69 | 3.71 | 1.28 | |||
表7 交吾托海野果林居群灌木生态位重叠指数Tab. 7 Shrub niche overlap index of JWTH wild fruit forest community |
| 物种 | 黑果小檗 Berberis atrocarpa | 小叶忍冬 Lonicera microphylla | 矮小忍冬 Lonicera humilis | 弯刺蔷薇 Rosa beggeriana | 疏花蔷薇 Rosa laxa |
|---|---|---|---|---|---|
| 小叶忍冬Lonicera microphylla | 0.22 | ||||
| 矮小忍冬Lonicera humilis | 0.52 | 0.02 | |||
| 弯刺蔷薇Rosa beggeriana | 0.26 | 0.00 | 0.43 | ||
| 疏花蔷薇Rosa laxa | 0.00 | 0.00 | 0.00 | 0.86 | |
| 樱桃李Prunus cerasifera | 0.37 | 0.00 | 0.00 | 0.00 | 0.00 |
表8 杏花沟野果林居群灌木生态位重叠指数Tab. 8 Shrub niche overlap index of XHG wild fruit forest community |
| 物种 | 小叶忍冬 Lonicera microphylla | 矮小忍冬 Lonicera humilis | 新疆忍冬 Lonicera tatarica | 疏花蔷薇 Rosa laxa | 沙棘 Hippophae rhamnoides |
|---|---|---|---|---|---|
| 矮小忍冬Lonicera humilis | 0.66 | ||||
| 新疆忍冬Lonicera tatarica | 0.00 | 0.00 | |||
| 疏花蔷薇Rosa laxa | 0.48 | 0.00 | 0.00 | ||
| 沙棘Hippophae rhamnoides | 0.57 | 0.31 | 0.00 | 0.71 | |
| 欧洲木莓Rubus caesius | 0.00 | 0.00 | 0.00 | 0.00 | 0.71 |
表9 莫乎尔野果林居群灌木生态位重叠指数Tab. 9 Shrub niche overlap index of MHE wild fruit forest community |
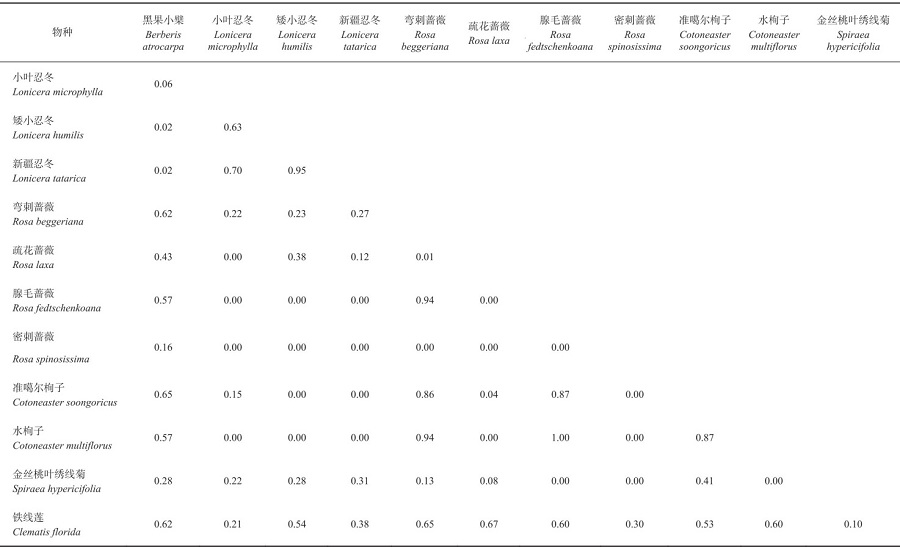 |
表10 大西沟野果林居群灌木生态位重叠指数Tab. 10 Shrub niche overlap index of DXG wild fruit forest community |
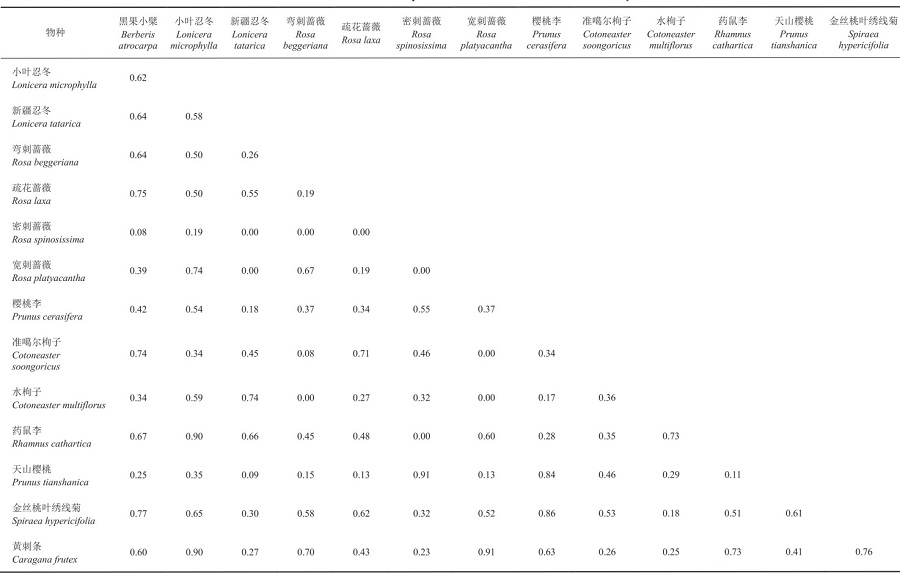 |
表11 4个野果林居群的灌木间总体联结性分析Tab. 11 Analysis of the overall correlation between shrubs in four wild fruit forest populations |
| 系数 | 方差比率(VR) | 检验统计量(W) | 临界值( , ) | 测度结果 | ||
|---|---|---|---|---|---|---|
| 交吾托海JWTH | 1.11 | 0.54 | 0.49 | 4.40 | (3.94,18.31) | 不显著负关联 |
| 杏花沟XHG | 0.94 | 1.11 | 1.18 | 10.66 | (3.94,18.31) | 不显著正关联 |
| 莫乎尔MHE | 2.00 | 1.21 | 0.61 | 5.44 | (3.94,18.31) | 不显著负关联 |
| 大西沟DXG | 2.27 | 2.32 | 1.02 | 9.20 | (3.94,18.31) | 不显著正关联 |
| [1] |
马克平. 试论生物多样性的概念[J]. 生物多样性, 1993, 1(1): 20-22.
[
|
| [2] |
马晓勇, 上官铁梁. 太岳山森林群落物种多样性[J]. 山地学报, 2004, 22(5): 606-612.
[
|
| [3] |
董雪, 辛智鸣, 段瑞兵, 等. 乌兰布和沙漠典型灌木群落多样性及其生态位[J]. 干旱区研究, 2020, 37(4): 1009-1017.
[
|
| [4] |
|
| [5] |
赵阳, 曹家豪, 李波, 等. 洮河自然保护区大峪沟林区紫果云杉群落木本植物种群生态位特征[J]. 生态学报, 2022, 42(5): 1865-1875.
[
|
| [6] |
|
| [7] |
陈铭, 黄林娟, 黄贵, 等. 广西大石围天坑群草本植物多样性及其生态位变化规律[J]. 生态学报, 2023, 43(7): 2831-2844.
[
|
| [8] |
于飞, 王洋, 张岗岗, 等. 游荡型黄河滩涂植物群落多样性及种间联结性[J]. 生态学报, 2023, 43(6): 2429-2439.
[
|
| [9] |
高金辉, 刘运伟, 韩家永, 等. 小兴安岭刺五加群落植物组成及区系分析[J]. 森林工程, 2021, 37(6): 39-46.
[
|
| [10] |
曹晓栋, 杨波, 黄梅, 等. 贵州省宽阔水国家级自然保护区草本植物区系及物种多样性研究[J]. 西北植物学报, 2021, 41(9): 1559-1569.
[
|
| [11] |
褚佳瑶, 冯琳骄, 侯毅兴, 等. 新疆野苹果种群受损现状[J]. 经济林研究, 2022, 40(1): 265-273.
[
|
| [12] |
翟朝阳, 邱娟, 司洪章, 等. 微地形对大西沟新疆野杏萌发层土壤因子的影响[J]. 生态学报, 2019, 39(6): 2168-2179.
[
|
| [13] |
张小燕, 梅闯, 闫鹏, 等. 新疆野苹果枝条解剖结构与苹果小吉丁虫抗性的关系[J]. 新疆农业科学, 2021, 58(3): 540-546.
[
|
| [14] |
李利平, 海鹰, 安尼瓦尔·买买提, 等. 新疆伊犁地区野果林的群落特征及保护[J]. 干旱区研究, 2011, 28(1): 60-66.
[
|
| [15] |
王超, 周龙, 彭妮, 等. 天山樱桃花芽形态分化研究[J]. 西北农业学报, 2015, 24(5): 97-101.
[
|
| [16] |
李林瑜, 方紫妍, 艾克拜尔·毛拉, 等. 不同天然居群小檗种子萌发障碍因子研究[J]. 植物研究, 2018, 38(6): 894-901.
[
|
| [17] |
林培钧, 崔乃然. 天山野果林资源——伊犁野果林综合研究[M]. 北京: 中国林业出版社, 2000: 76-82.
[
|
| [18] |
侯博, 许正. 中国伊犁野生果树及近缘种研究[J]. 干旱区研究, 2006, 23(3): 453-458.
[
|
| [19] |
方紫妍, 李林瑜, 艾克拜尔·毛拉, 等. 新疆西天山土壤生境质量与野果林群落特征的相关性分析[J]. 福建农林大学学报(自然科学版), 2019, 48(4): 453-458.
[
|
| [20] |
|
| [21] |
石荡, 郭传超, 蒋南林, 等. 新疆野杏天然更新幼株的个体特征及空间分布格局[J]. 植物生态学报, 2023, 47(4): 515-529.
[
|
| [22] |
吴征镒. 《世界种子植物科的分布区类型系统》的修订[J]. 云南植物研究, 2003, 25(5): 535-538.
[
|
| [23] |
吴征镒, 周浙昆, 孙航, 等. 种子植物分布区类型及其起源和分化[M]. 昆明: 云南科学技术出版社, 2006: 146-451.
[
|
| [24] |
王国宏, 方精云, 郭柯, 等. 《中国植被志》研编内容与规范[J]. 植物生态学报, 2020, 44(2): 128-178.
[
|
| [25] |
布海丽且姆·阿卜杜热合曼, 刘会良, 刘娜, 等. 天山野果林种子植物区系分析[J]. 干旱区地理, 2016, 39(4): 828-834.
[
|
| [26] |
崔大方, 廖文波, 羊海军, 等. 中国伊犁天山野果林区系表征地理成分及区系发生的研究[J]. 林业科学研究, 2006, 19(5): 555-560.
[
|
| [27] |
黄睿智, 王奇, 孙婧依, 等. 太白山南北坡栎类林物种组成与群落特征比较[J]. 应用生态学报, 2023, 34(8): 2055-2064.
[
|
| [28] |
侯予红. 新疆小檗属植物的识别及生物学特性[J]. 防护林科技, 2018(12): 69, 79.
[
|
| [29] |
杨星, 史洁婷, 许文超, 等. 黑龙江省高速公路中央分隔带植物选择与配置[J]. 北方园艺, 2015(7): 72-76.
[
|
| [30] |
刘英. 西安不同类型城市绿地中园林植物应用研究[D]. 杨凌: 西北农林科技大学, 2020.
[
|
| [31] |
郭香瑶, 罗颖, 尹秋龙, 等. 秦岭皇冠暖温带落叶阔叶林灌木层结构与物种多样性[J]. 应用生态学报, 2022, 33(8): 2017-2026.
[
|
| [32] |
田蕾, 周璐, 孙佳瑞, 等. 山地乔灌木斑块空间分布格局及其多样性沿海拔梯度的变化规律[J]. 生态学报, 2023, 43(24): 10320-10333.
[
|
| [33] |
王继丰, 韩大勇, 王建波, 等. 三江平原湿地小叶章群落沿土壤水分梯度物种组成及多样性变化[J]. 生态学报, 2017, 37(10): 3515-3524.
[
|
| [34] |
高军, 杨建英, 史常青, 等. 密云水库上游油松人工水源涵养林林下植物多样性与土壤理化特性[J]. 应用生态学报, 2022, 33(9): 2305-2313.
[
|
| [35] |
宋兆斌, 罗凤敏, 马媛, 等. 乌兰布和—西鄂尔多斯荒漠草原过渡带典型灌木群落及其多样性[J]. 水土保持通报, 2022, 42(5): 65-72.
[
|
| [36] |
黄林娟, 于燕妹, 安小菲, 等. 天坑森林植物群落叶功能性状、物种多样性和功能多样性特征[J]. 生态学报, 2022, 42(24): 10264-10275.
[
|
| [37] |
喻阳华, 钟欣平, 郑维, 等. 喀斯特森林不同演替阶段植物群落物种多样性、功能性状、化学计量及其关联[J]. 生态学报, 2021, 41(6): 2408-2417.
[
|
| [38] |
杨小平, 王海鸥, 蒋丽丽, 等. 伊犁地区不同新疆野苹果种群的年龄结构分析[J]. 分子植物育种, 2021, 19(12): 4133-4141.
[
|
| [39] |
|
| [40] |
李帅锋, 刘万德, 苏建荣, 等. 滇西北云南红豆杉群落物种生态位与种间联结[J]. 植物科学学报, 2012, 30(6): 568-576.
[
|
| [41] |
|
| [42] |
黄睿雪, 欧静, 王洪飞, 等. 百里杜鹃国家森林公园植物群落物种多样性及优势种生态位宽度对人为干扰的响应[J]. 生态学杂志, 2024, 43(5): 1217-1226.
[
|
| [43] |
钟欣艺, 赵凡, 姚雪, 等. 三星堆遗址城墙不同维护措施下草本植物群落组成、生态位及稳定性特征[J]. 应用生态学报, 2023, 34(11): 2938-2946.
[
|
| [44] |
牛慧慧, 陈辉, 付阳, 等. 柴达木盆地东部荒漠植物生态位特征[J]. 生态学报, 2019, 39(8): 2862-2871.
[
|
| [45] |
|
| [46] |
陈聪琳, 赵常明, 刘明伟, 等. 神农架南坡小叶青冈+曼青冈常绿阔叶林主要木本植物生态位与种间联结[J]. 生态学报, 2024, 44(11): 4889-4903.
[
|
| [47] |
钱逸凡, 伊力塔, 胡军飞, 等. 普陀山主要植物种生态位特征[J]. 生态学杂志, 2012, 31(3): 561-568.
[
|
| [48] |
陈丝露, 赵敏, 李贤伟, 等. 柏木低效林不同改造模式优势草本植物多样性及其生态位[J]. 生态学报, 2018, 38(1): 143-155.
[
|
| [49] |
庞久帅, 原阳晨, 周苗苗, 等. 槭叶铁线莲(Clematis acerifolia Maxim.)群落物种生态位研究[J]. 生态学报, 2022, 42(8): 3449-3457.
[
|
| [50] |
陈玉凯, 杨琦, 莫燕妮, 等. 海南岛霸王岭国家重点保护植物的生态位研究[J]. 植物生态学报, 2014, 38(6): 576-584.
[
|
| [51] |
刘润红, 陈乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J]. 生态学报, 2020, 40(6): 2057-2071.
[
|
| [52] |
涂洪润, 农娟丽, 朱军, 等. 桂林岩溶石山密花树群落主要物种的种间关联及群落稳定性[J]. 生态学报, 2022, 42(9): 3688-3705.
[
|
| [53] |
张盟, 郑俊鸣, 万佳艺, 等. 福建省东门屿优势木本植物的生态位与种间联结[J]. 森林与环境学报, 2022, 42(1): 11-19.
[
|
| [54] |
蒋南林, 邱娟, 石荡, 等. 基于草本群落、牧道特征对野果林放牧干扰强度的评价[J]. 生态学报, 2024, 44(7): 3060-3071.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}