
Effects of different habitats and substrates on seed germination and seedling growth of Malus sieversii after passing through digestive tract of cattle
Received date: 2023-11-01
Revised date: 2024-03-12
Online published: 2024-06-20
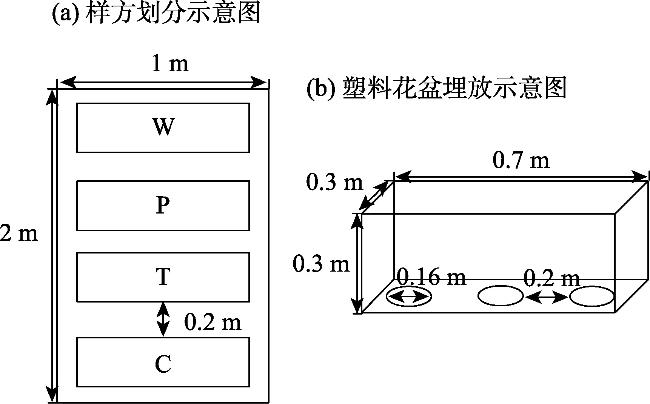
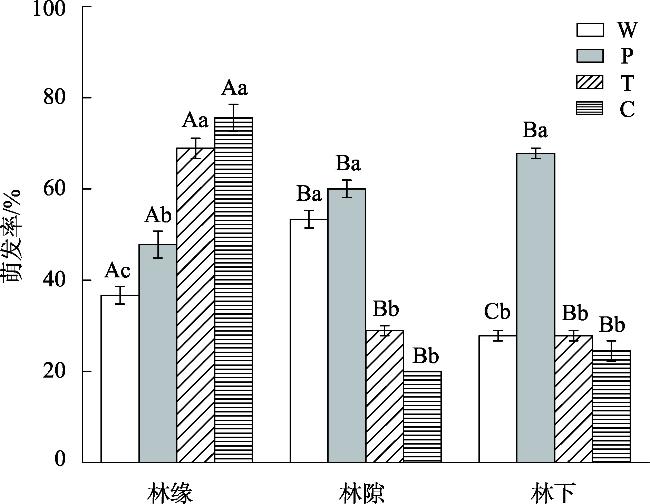
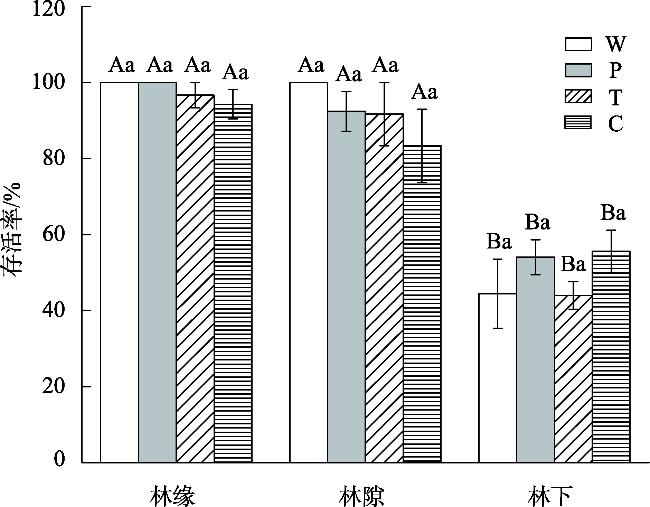
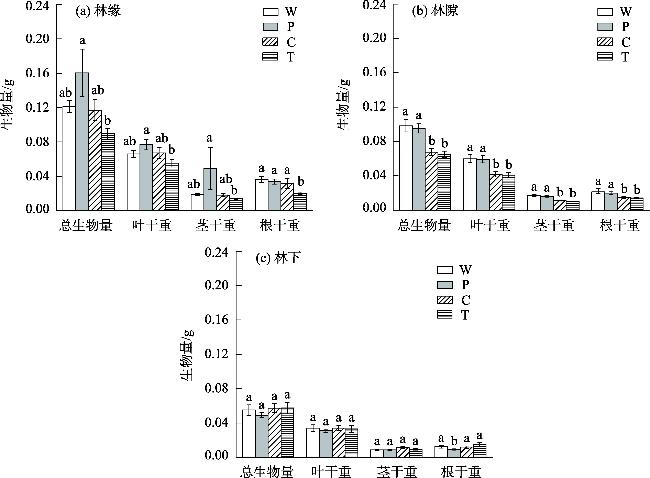
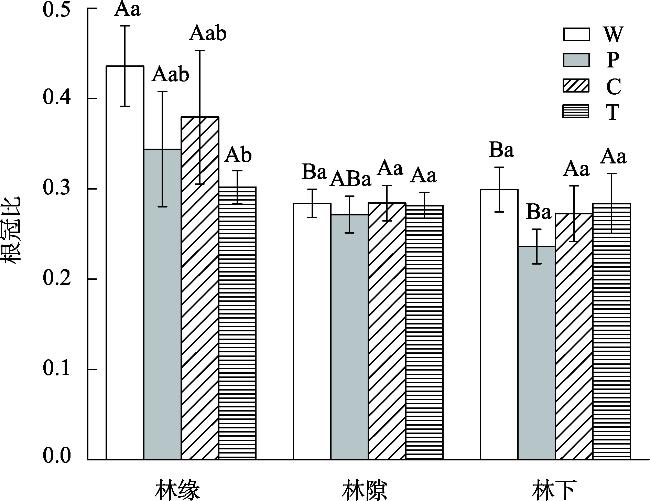
This study aimed to reveal the effects of different habitats and substrates on seed germination and seedling growth of Malus sieversii after passing through digestive tract of cattle. A controlled experiment was set up in field based on the different habitats and substrates in which seeds were deposited after passing through the digestive tract of cattle. The three habitats were as follows: forest edges, gaps, and understory. The four treatments were as follows: W (seeds through digestive tract+intact feces), P (seeds through digestive tract+broken feces), T (seeds through digestive tract+soil), and control C (seeds not through digestive tract+soil). The results showed that: The seed germination rates, seedling survival rate, and seedling biomass of M. sieversii were significantly different (P<0.05) in different habitats and treatments. In the forest edge habitat, seeds germinated rate was higher in soil substrate compared to fecal substrate, while the gaps and understory habitats showed the opposite trend. Seedling survival rate was significantly higher in the forest edge and gaps habitats compared to the understory habitat. Under the same treatment, seedling biomass was higher in the forest edge compared to the gaps, and higher in the gaps compared to the understory. Within the same habitat, seedling biomass from fecal substrate in forest edge and gap habitats surpassed that of soil substrate. The study revealed that the heterogeneity of habitat and substrate is the main reason that affects the seed germination and seedling establishment of M. sieversii after passing through digestive tract of cattle.
Key words: Malus sieversii; endozoochory; habitats; substrate; seed germination; seedling growth
BAI Shilin , LYU Yaya , SHI Xiaojun . Effects of different habitats and substrates on seed germination and seedling growth of Malus sieversii after passing through digestive tract of cattle[J]. Arid Zone Research, 2024 , 41(5) : 821 -829 . DOI: 10.13866/j.azr.2024.05.10
表1 不同生境和处理下新疆野苹果幼苗的生长特征Tab. 1 Growth characteristics of Malus sieversii seedlings in different habitats and treatments |
| 生境 | 处理 | 株高/mm | 基茎/mm | 主根长/mm | 叶片数/个 |
|---|---|---|---|---|---|
| 林缘 | W | 43.99±0.83Aa | 1.47±0.03Ab | 90.98±4.18Aa | 6.00±0.20Aa |
| P | 41.24±1.29Aa | 1.61±0.03Aa | 85.76±3.27Aa | 5.80±0.20Aab | |
| T | 34.87±1.20Ab | 1.41±0.03Ab | 72.87±9.64Ab | 6.00±0.14Aa | |
| C | 34.76±1.10Ab | 1.25±0.02Ac | 79.32±2.54Aab | 5.40±0.19Ab | |
| 林隙 | W | 51.30±1.39Aa | 1.42±0.04Aa | 75.10±3.95ABa | 5.20±0.17ABa |
| P | 43.51±1.13Ab | 1.46±0.03Bab | 71.81±3.17Ba | 5.671±0.16Aa | |
| T | 32.24±1.39Ac | 1.39±0.03Aab | 83.06±5.78Aa | 5.33±0.13Aa | |
| C | 28.67±0.70Bc | 1.30±0.03Ab | 84.72±7.60Aa | 5.56±0.18Aa | |
| 林下 | W | 38.54±2.59Ba | 1.20±0.02Bb | 65.13±4.96Ba | 4.18±0.28Ba |
| P | 34.06±1.25Bab | 1.21±0.02Cb | 66.18±1.49Ba | 4.20±0.14Ba | |
| T | 29.96±1.03Aab | 1.30±0.02Aa | 69.49±1.63Aa | 4.33±0.24Ba | |
| C | 27.64±1.35Bb | 1.29±0.02Aa | 71.59±2.74Aa | 3.80±0.24Ba |
注:不同大写字母表示相同处理不同生境间存在显著差异(P<0.05),不同小写字母表示同一生境不同处理间存在显著差异(P<0.05)。 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
王树林, 侯扶江. 粪种子库的理论基础、影响因素和生态意义[J]. 生态学报, 2023, 43(11): 4369-4389.
[
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
褚佳瑶, 冯琳骄, 侯毅兴, 等. 新疆野苹果种群受损现状[J]. 经济林研究, 2022, 40(1): 265-273.
[
|
| [17] |
米尔卡米力·麦麦提, 刘忠权, 马晓东, 等. 新疆野苹果的生存现状、问题及保护策略[J]. 广西植物, 2021, 41(12): 2100-2109.
[
|
| [18] |
|
| [19] |
许宁, 憨宏艳, 甘小洪. 光照及地面覆盖物对水青树种子萌发和幼苗初期生长的影响[J]. 植物资源与环境学报, 2015, 24(3): 85-93.
[
|
| [20] |
谭美, 杨志玲, 杨旭, 等. 不同生境内厚朴种子萌发和幼苗生长研究[J]. 生态与农村环境学报, 2018, 34(10): 910-916.
[
|
| [21] |
张明, 芦光新, 王伟, 等. 牦牛粪便对燕麦(Avena sativa)种子发芽及幼苗生长的影响[J]. 黑龙江畜牧兽医, 2014(11): 103-105.
[
|
| [22] |
高贤明, 杜晓军, 王中磊. 北京东灵山区两种生境条件下辽东栎幼苗补充与建立的比较[J]. 植物生态学报, 2003, 27(3): 404-411.
[
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
外里娜丽克斯, 王树林, 赵刚, 等. 4种豆科牧草种子对绵羊消化道作用的反应[J]. 草业科学, 2016, 33(8): 1566-1573.
[
|
| [27] |
张宗芳, 徐将, 师小军. 新疆野苹果幼苗生长及生物量分配对降水量和降水间隔时间的响应[J]. 干旱区研究, 2023, 40(1): 102-110.
[
|
| [28] |
吴泽民, 黄成林, 韦朝领. 黄山松群落林隙光能效应与黄山松的更新[J]. 应用生态学报, 2000, 11(1): 13-18.
[
|
| [29] |
张宗芳, 徐将, 师小军. 不同光照强度对新疆野苹果种子萌发和幼苗生长的影响[J]. 新疆农业大学学报, 2021, 44(6): 401-406.
[
|
| [30] |
徐来仙, 姚兰, 艾训儒, 等. 植被和凋落物对水杉天然幼苗萌发的影响[J]. 森林与环境学报, 2022, 42(5): 456-464.
[
|
| [31] |
巴音达拉, 江到无列提·米山别克. 新疆野苹果幼苗生存的距离制约效应研究[J]. 湖南农业科学, 2019(11): 75-77.
[Bayandala,
|
| [32] |
许中旗, 黄选瑞, 徐成立, 等. 光照条件对蒙古栎幼苗生长及形态特征的影响[J]. 生态学报, 2009, 29(3): 1121-1128.
[
|
| [33] |
|
| [34] |
华兆晖, 陶冶, 闫景明, 等. 氮添加对新疆野苹果幼苗枝叶大小的影响[J]. 干旱区研究, 2022, 39(1): 210-219.
[
|
| [35] |
任健, 李永进, 欧阳青, 等. 短期封育和施用牛粪对亚高山草甸植物群落组成及生物量的影响[J]. 草地学报, 2016, 24(6): 1197-1202.
[
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}