
Research progress on the mechanism of formation, absorption and utilization of condensed water on leaf trichomes of desert plants
Received date: 2023-06-11
Revised date: 2023-10-07
Online published: 2024-06-20
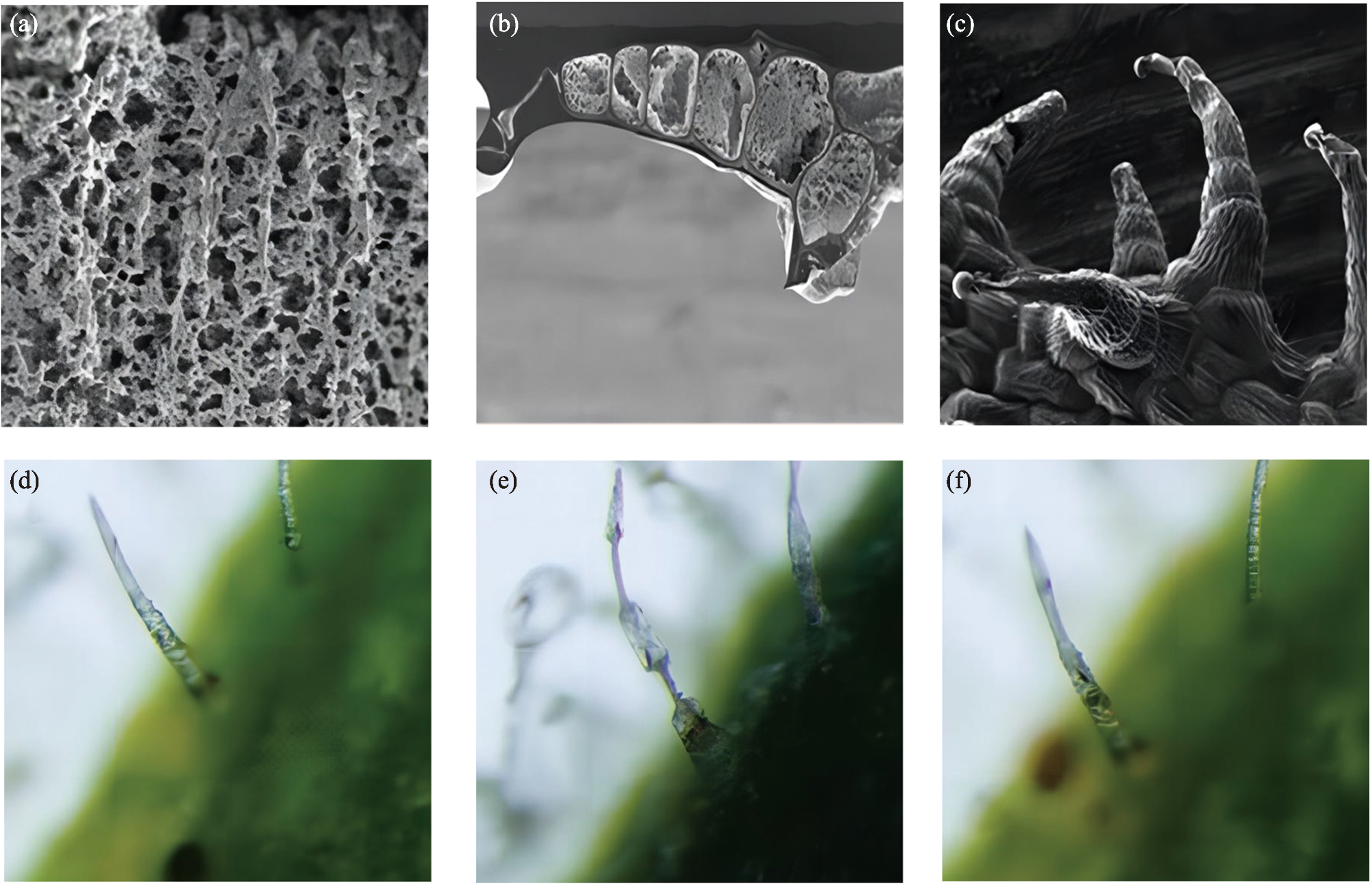
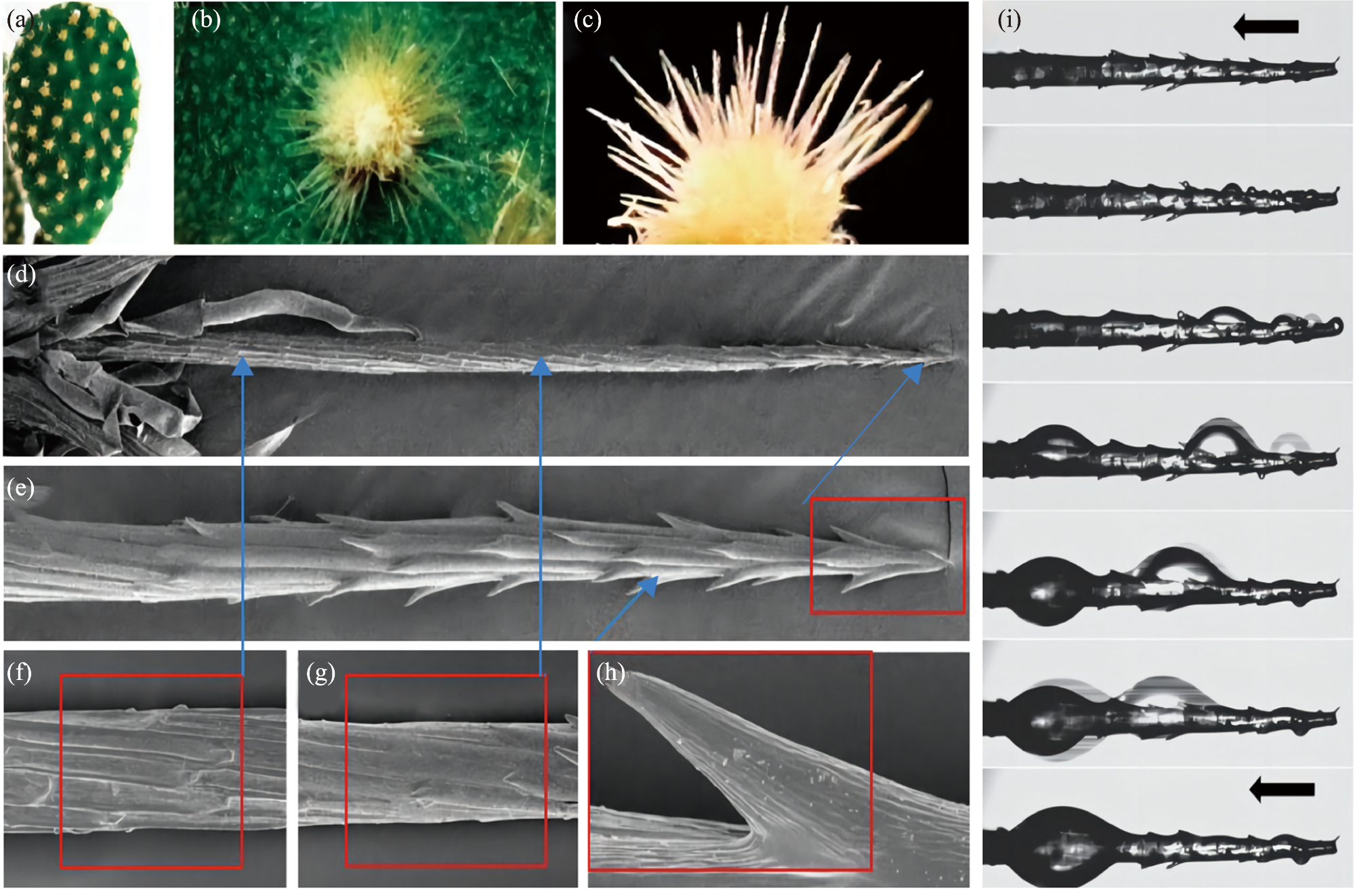
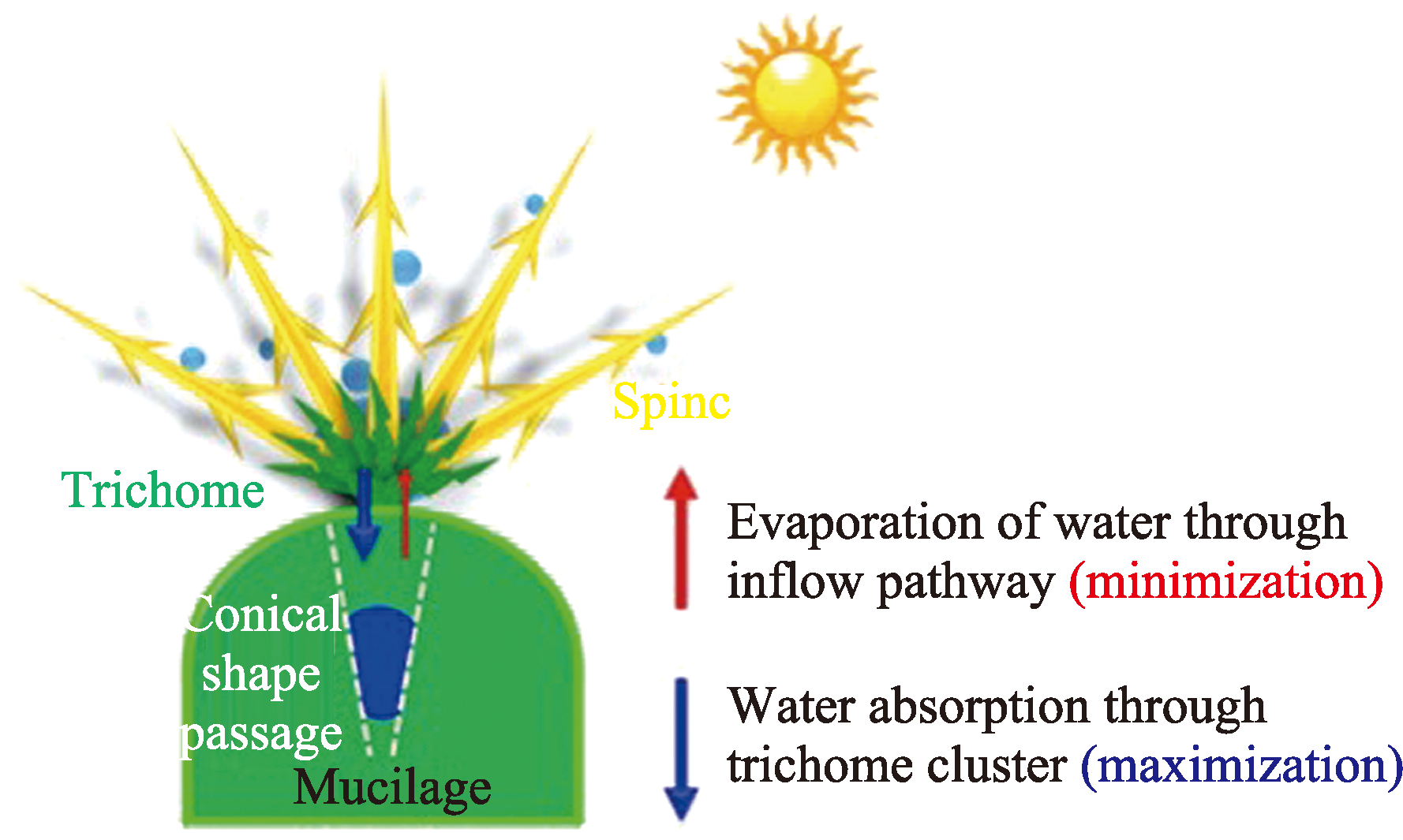
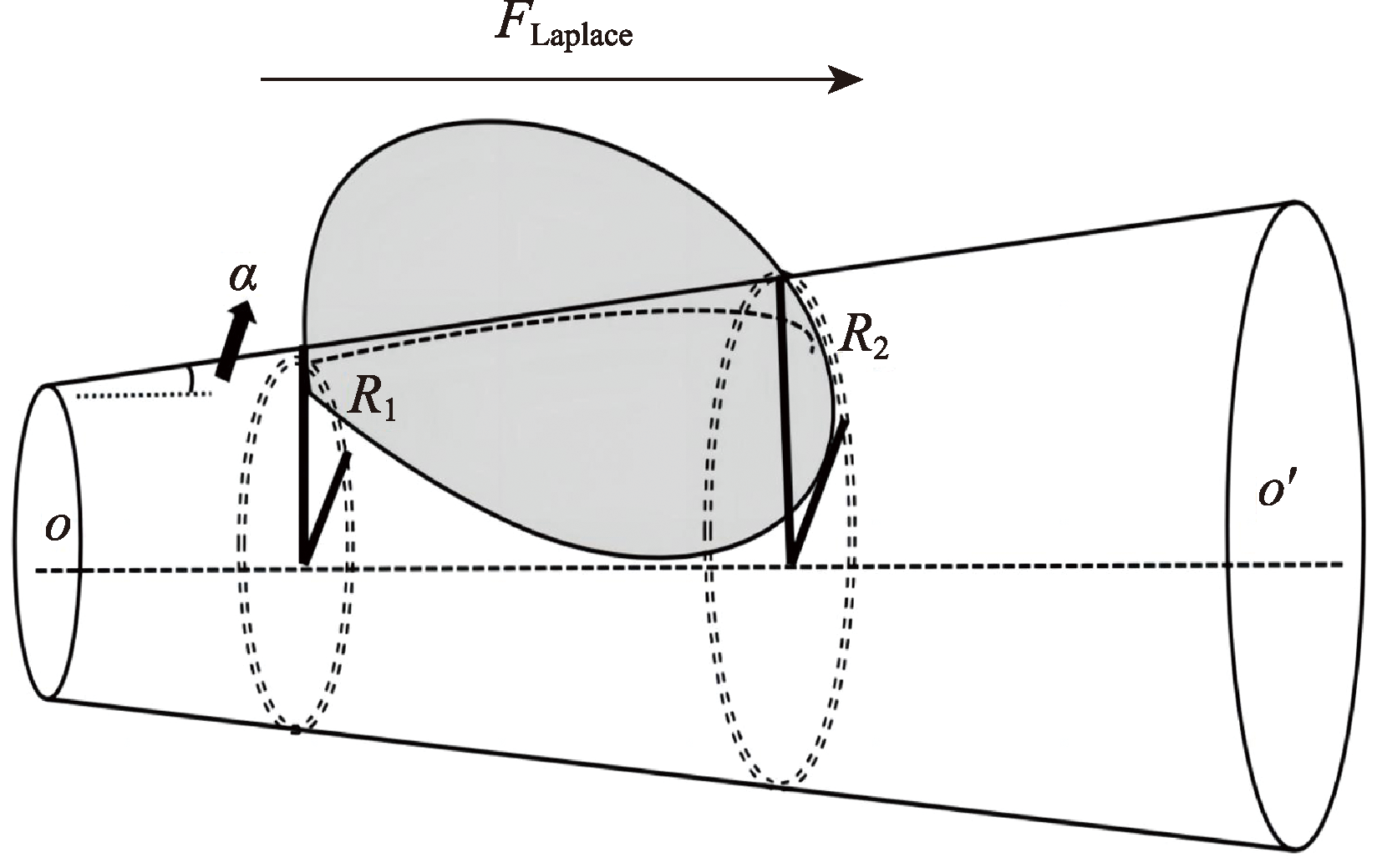
Desert plants have developed highly evolved water preservation and transportation systems for collecting and storing water through air under chronic water scarcity, the accessory structures such as trichomes on the leaf surface have important biological significance in collection, storage and transportation of the condensed water. Based on the current research progress, this review briefly summarizes the biological and ecological effects of condensate on desert plants, and expounds the theories related to leaf surface wettability and water collection capacity. Based on this, the important role of special structure of desert plants’ leaf surface (trichome) in the formation, transport and absorption of condensate is mainly discussed. The research methods of surface retention and absorption of condensate water were summarized, which may provide guidance for further study on trichome function and is helpful to understand the mechanism of condensate collection by desert plants driven by its structural characteristics in arid areas of China.
Key words: desert plant; condensed water; leaf surface wettability; trichome; ecological effect
Alayi HANATI , LIU Yanxia , LAN Haiyan . Research progress on the mechanism of formation, absorption and utilization of condensed water on leaf trichomes of desert plants[J]. Arid Zone Research, 2024 , 41(1) : 114 -123 . DOI: 10.13866/j.azr.2024.01.11
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
庄艳丽, 赵文智. 荒漠植物雾冰藜和沙米叶片对凝结水响应的模拟实验[J]. 中国沙漠, 2010, 30(5): 1068-1074.
[
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
刘志东, 吕光辉, 张雪妮, 等. 叶片被毛对干旱区短命植物狭果鹤虱生理生态特征的影响[J]. 干旱区研究, 2017, 34(5): 1101-1108.
[
|
| [35] |
李洁, 张铭芳. 植物表皮毛发育调控的研究进展[J]. 北方园艺, 2020, 44(23): 133-139.
[
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
/
| 〈 |
|
〉 |
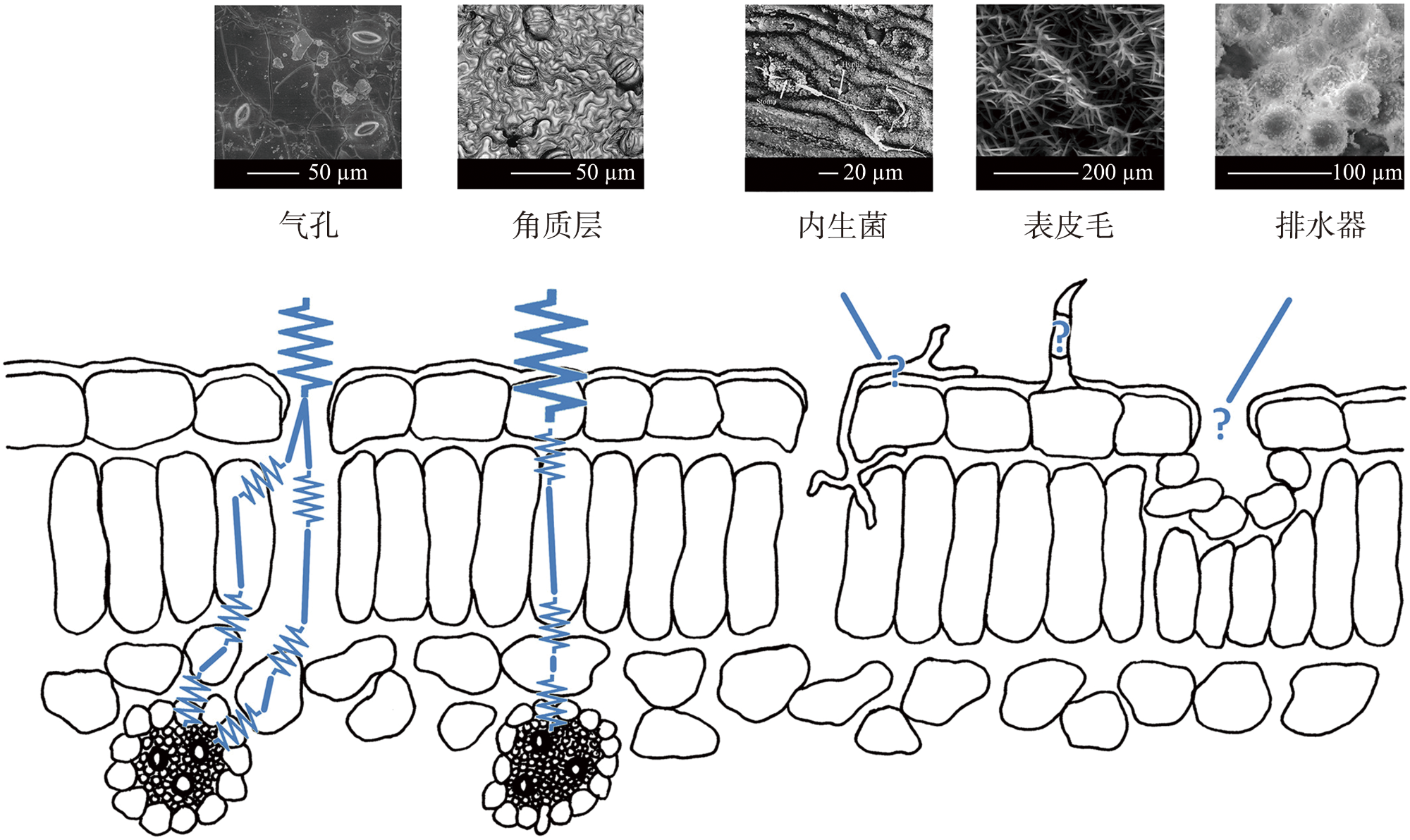
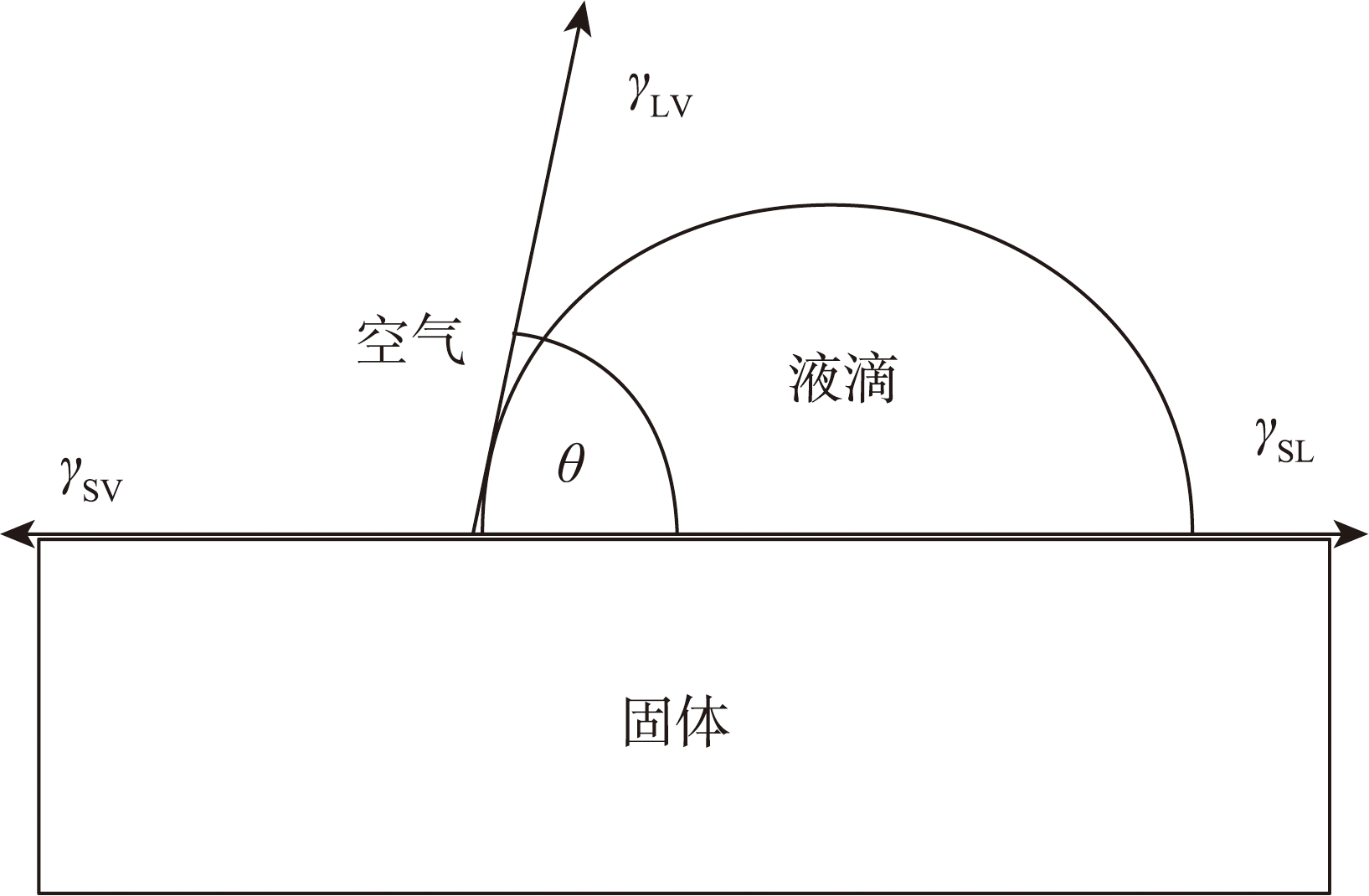
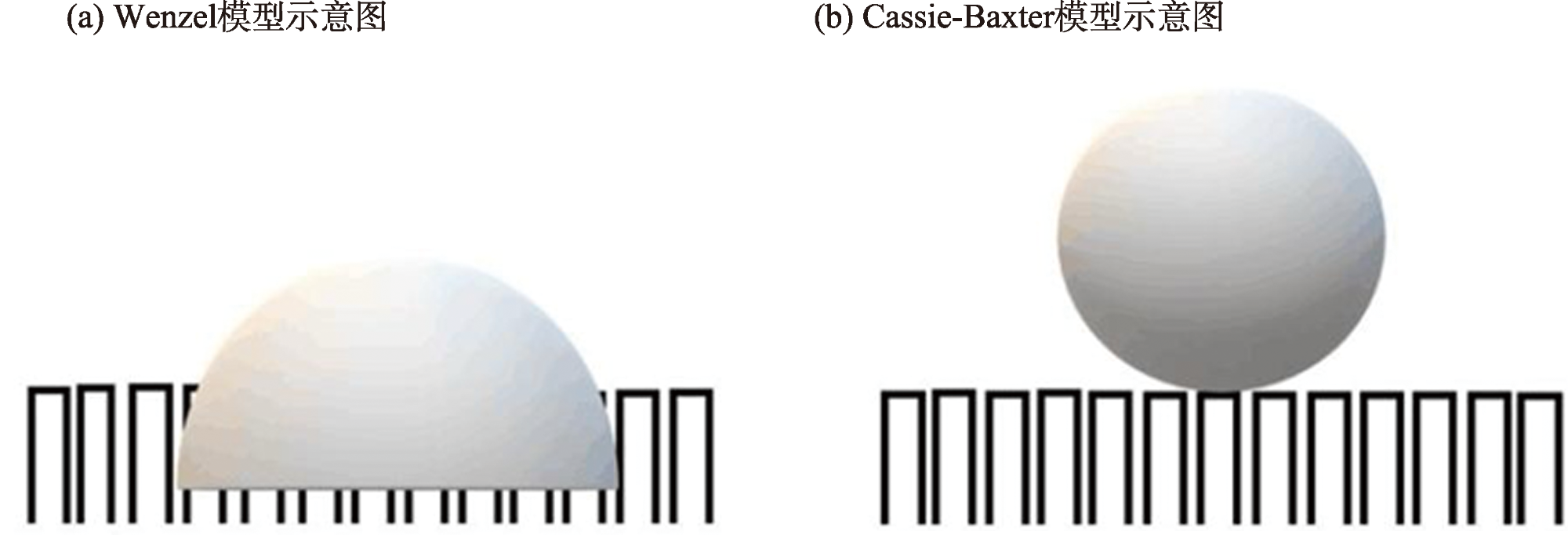
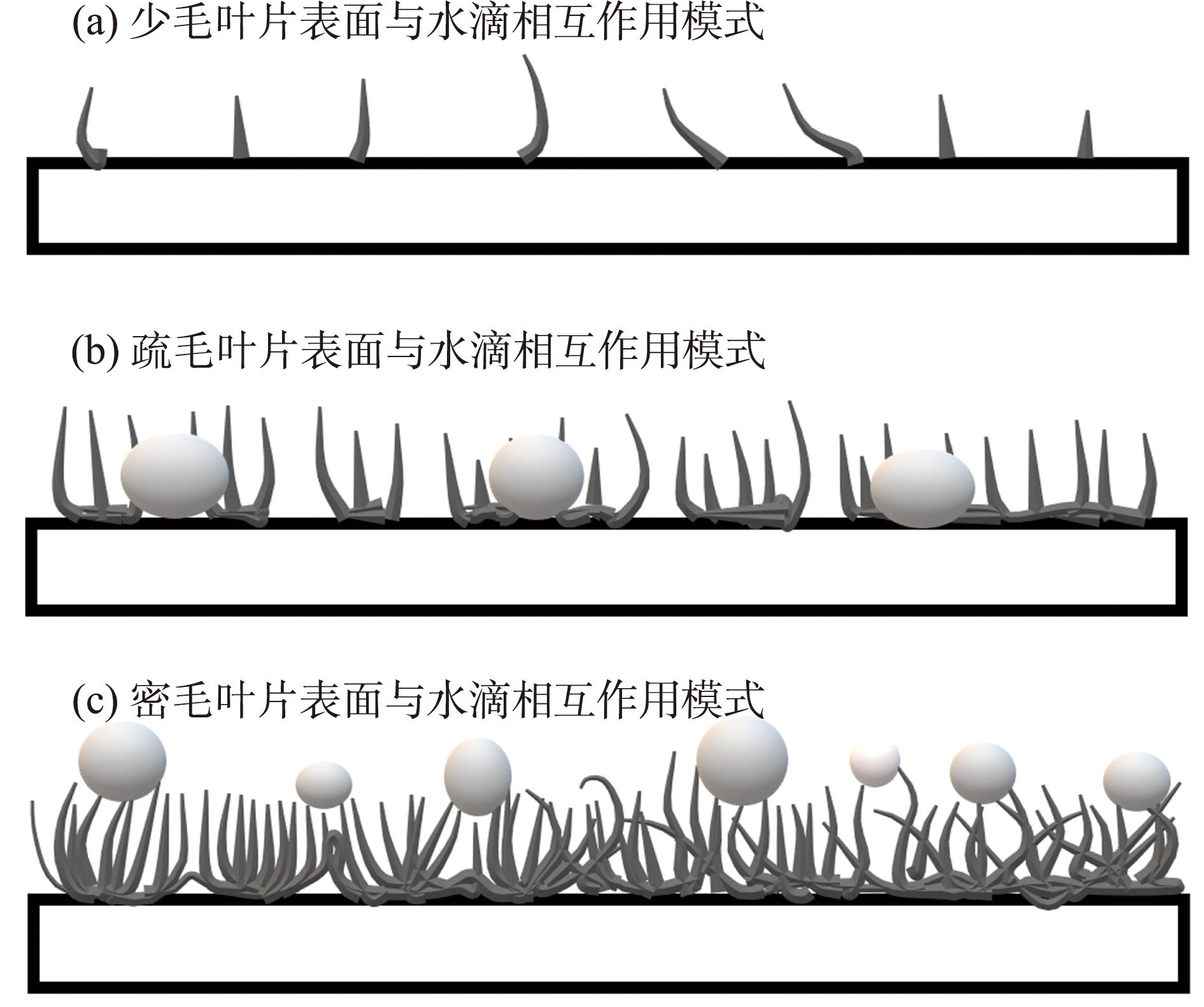
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}