
Effects of micro-topography on water use characteristics of alpine sand-fixing plants
Received date: 2023-07-11
Revised date: 2023-09-13
Online published: 2024-06-20
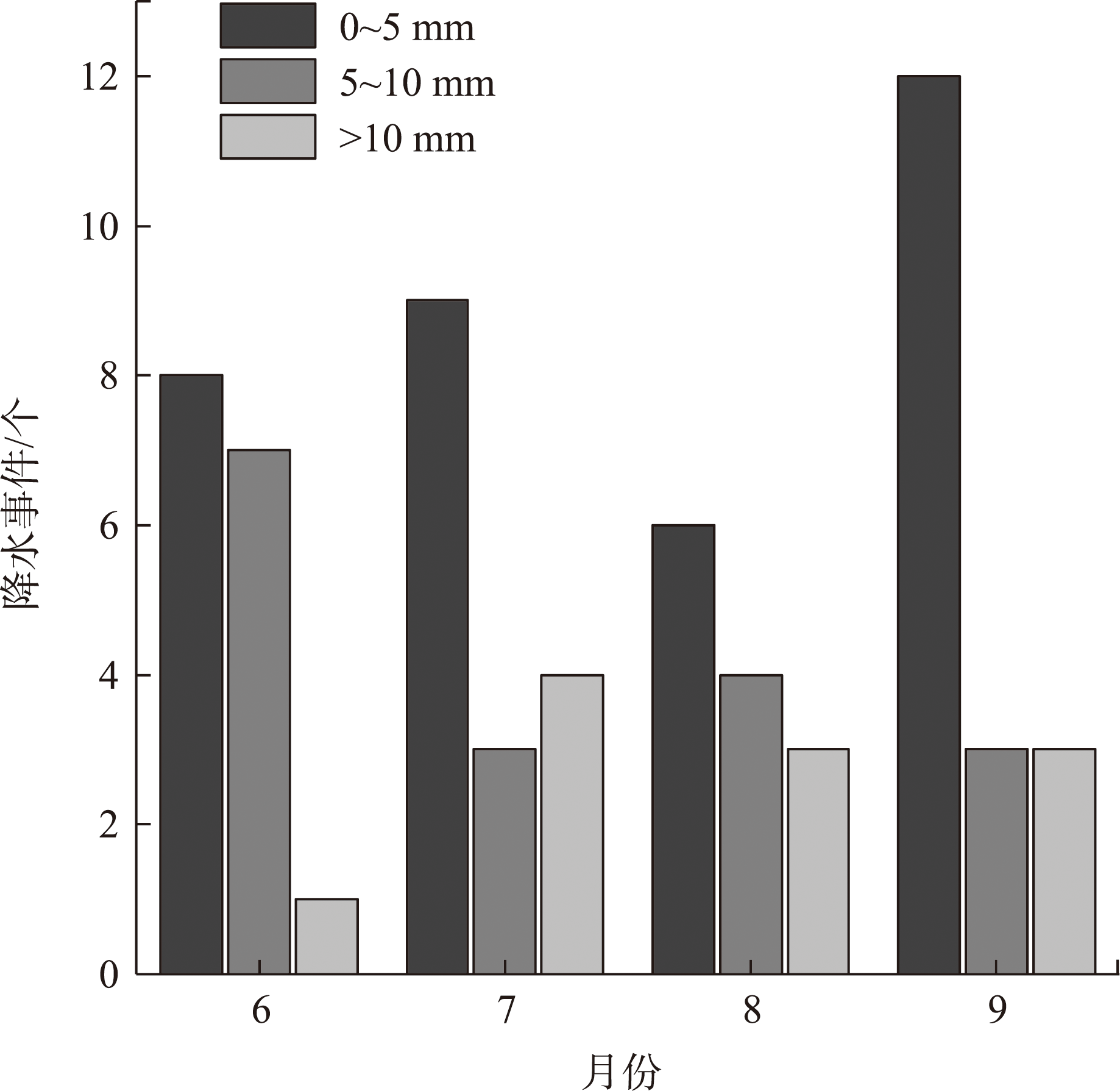
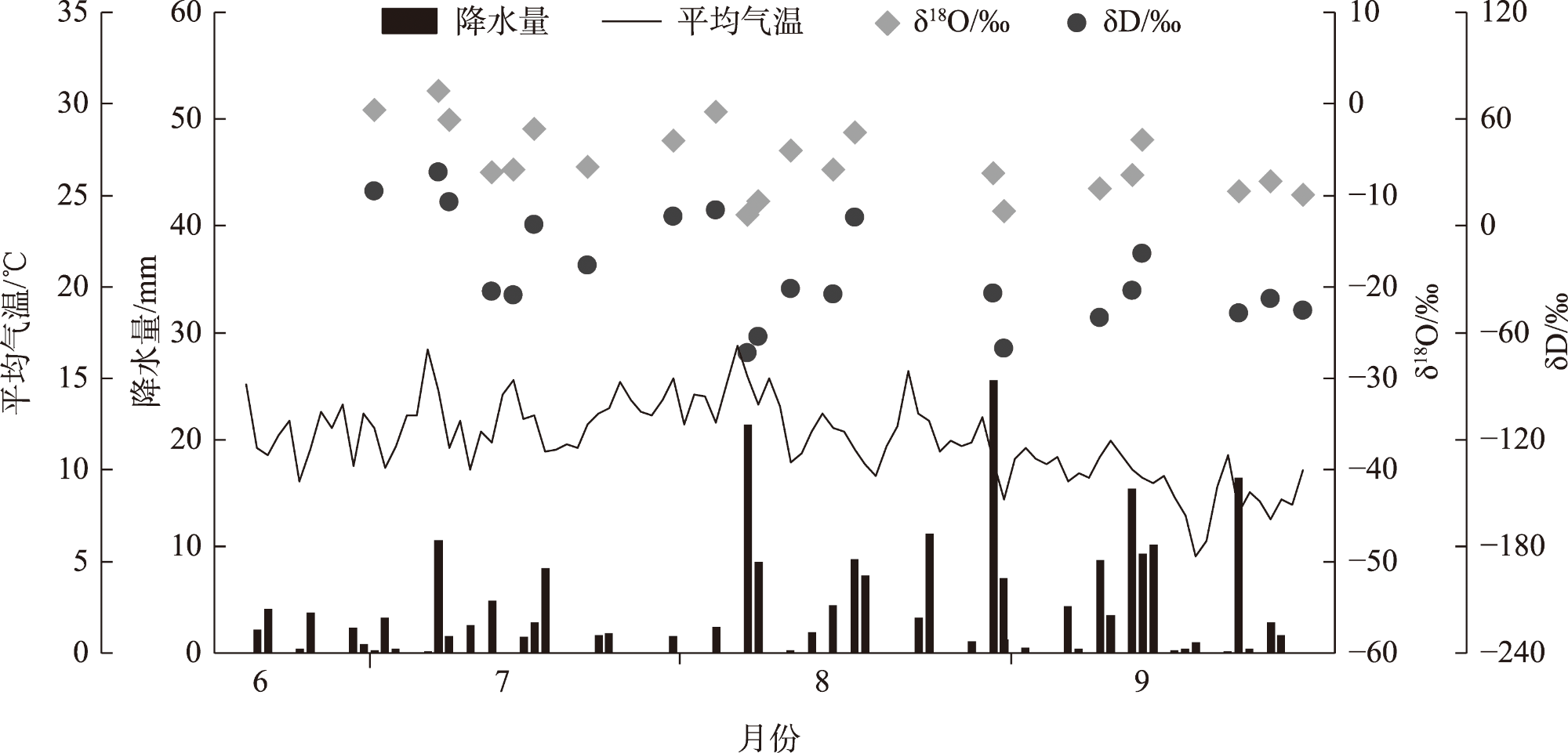
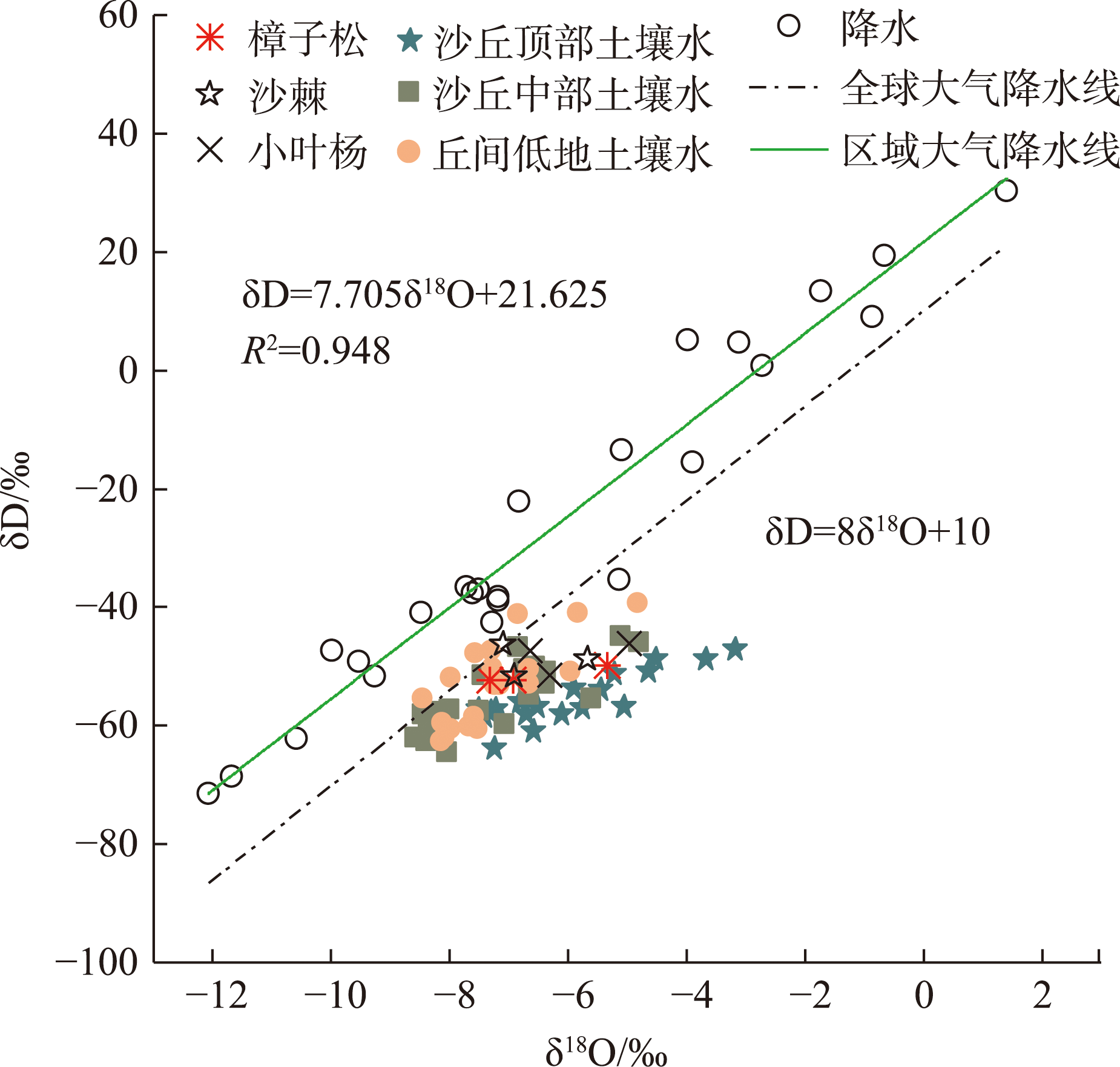
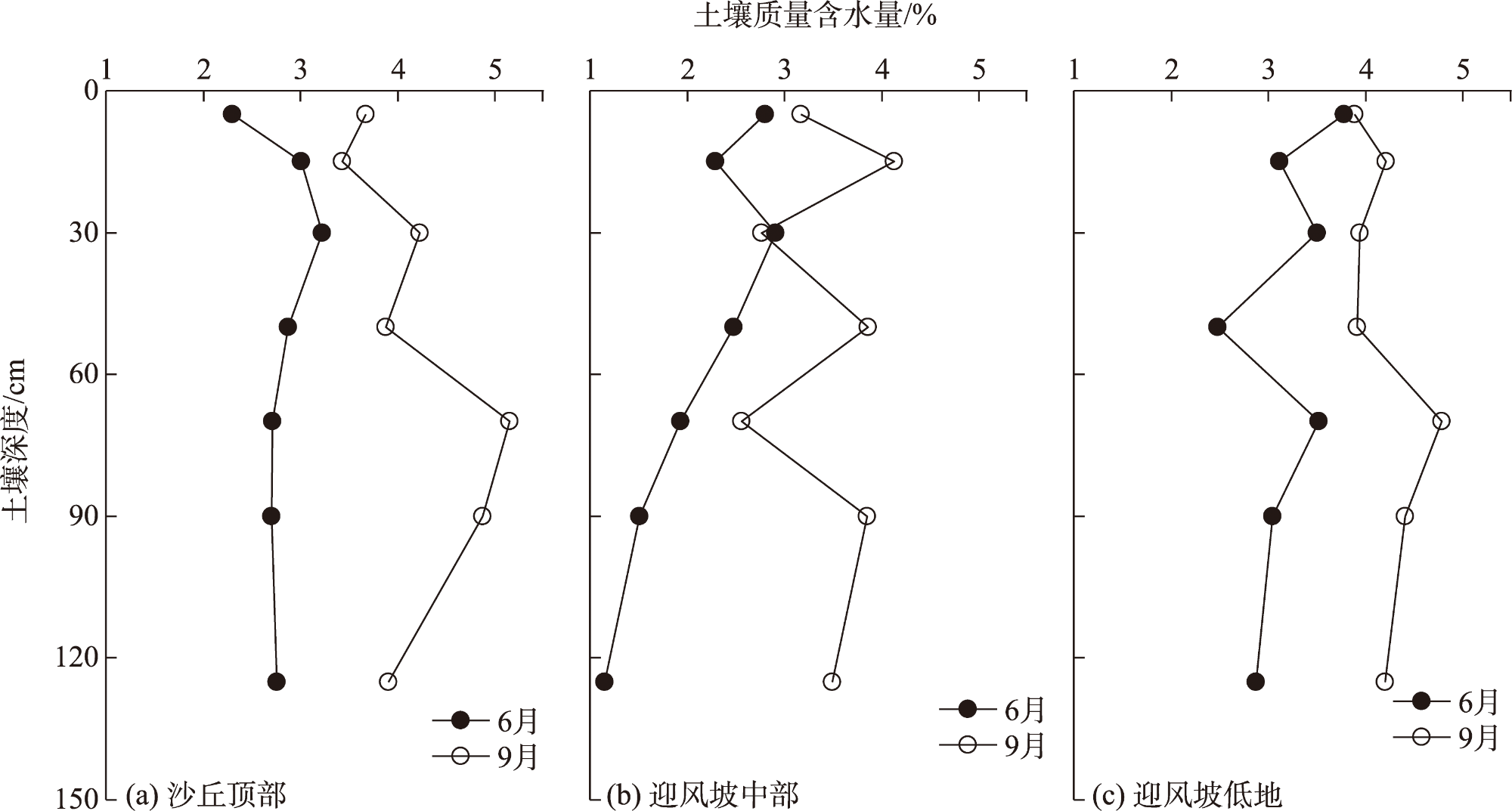
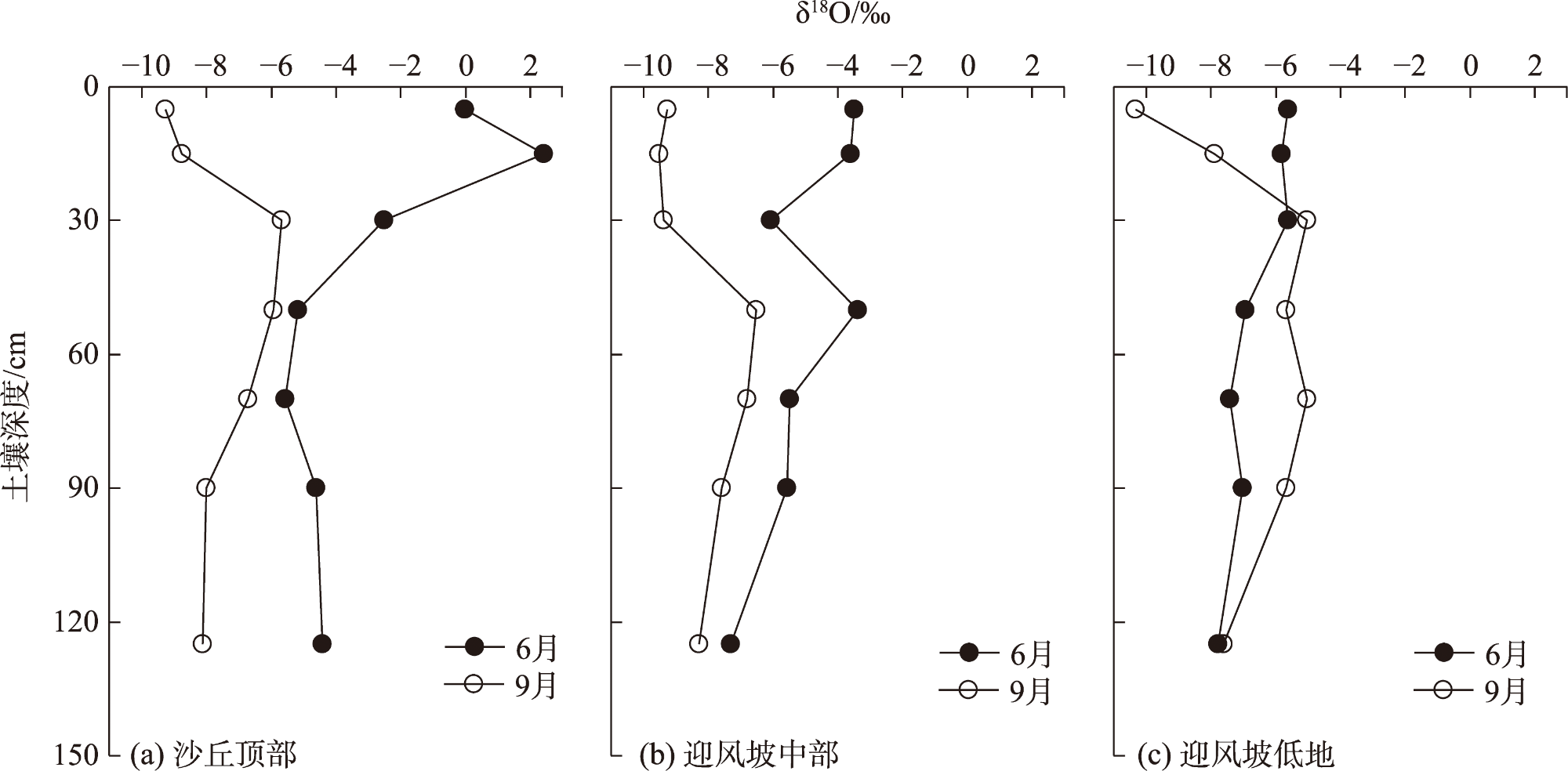
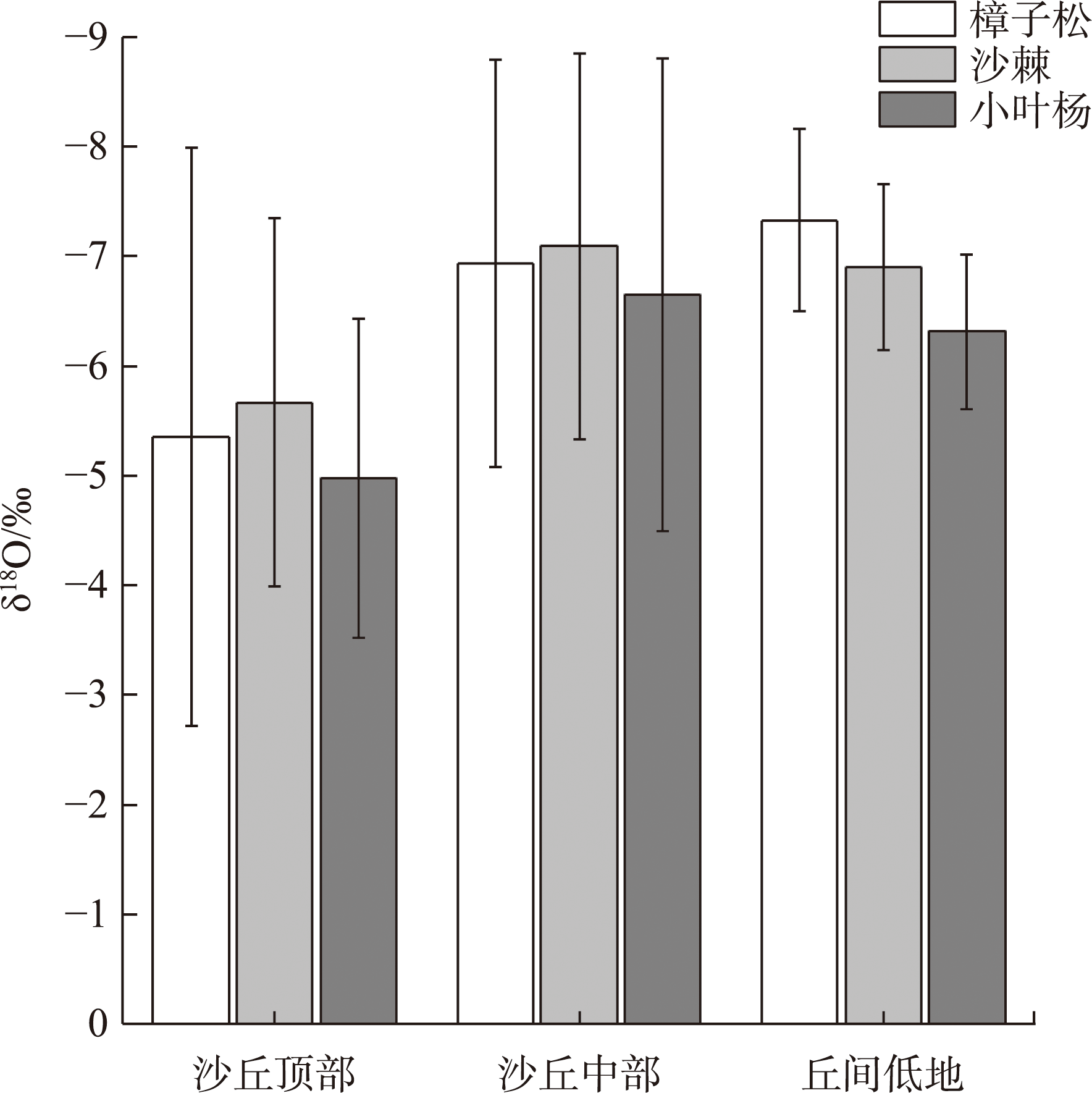
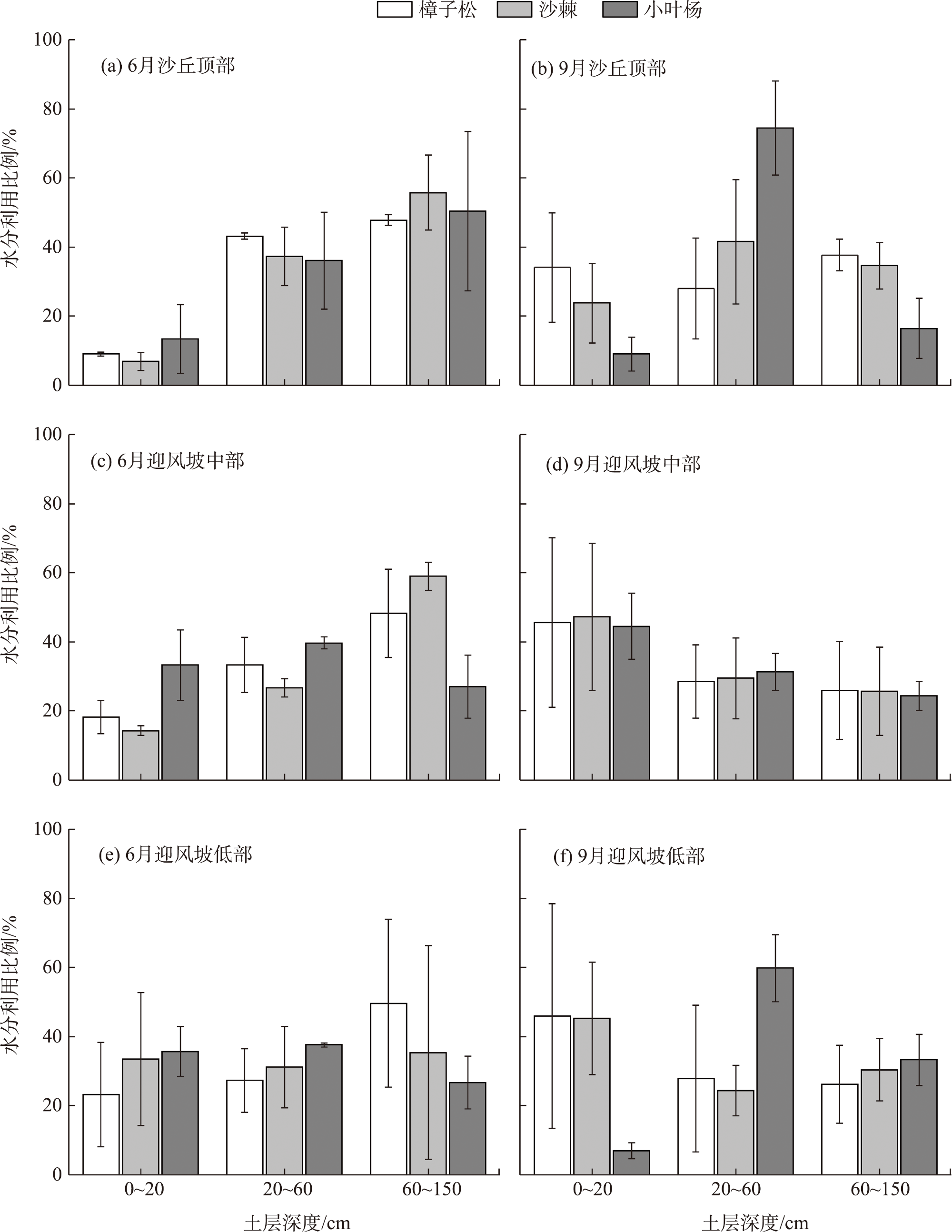
Water is the most critical limiting factor for plant survival, and the study of water utilization in desert plants has become the key to ecological protection and vegetation restoration programs. This study focused on three typical sand-fixing plants: Pinus sylvestris, Populus simonii, and Hippophae rhamnoides, in the sandy land on the east shore of Qinghai Lake, as the research material. The potential water source (varying levels of soil water) and the primary water sources for plants under the influence of micro-topography were analyzed by hydrogen and oxygen stable isotope technology (δ18O and δD) and the IsoSource isotope mixing model. The results show that: (1) the soil water content demonstrated micro-topography-based differences, which manifested as the soil water content on the windward slope being higher than that on the top of the sand dunes and the middle of the windward slope and the soil water content was at its highest in September. (2) the δ18O value of the xylem water varied in the tree species under different micro-topographic conditions. The δ18O values of P. sylvestris in the lowland of the windward slope were the lowest, while those of H. rhamnoides and P. simonii were the least in the middle of the windward slope. (3) marked seasonal variations were observed in the primary sources of water for different plants. In June, P. sylvestris and H. rhamnoides used deep soil water as the major source under varied micro-topographic conditions, while P. simonii mainly used deep soil water at the top of sand dunes. The soil water of the middle-layer was utilized more in the middle and lowlands of the windward slope. Still, with the increase of precipitation, various tree species turned to mainly using the shallow and middle soil water in September. In summary, the water use patterns of sand-fixing plants in the alpine sandy land were influenced by micro-topographic conditions, and varying species showed different degrees of response to precipitation.
FAN Mingyan , TIAN Lihui , ZHOU Hai . Effects of micro-topography on water use characteristics of alpine sand-fixing plants[J]. Arid Zone Research, 2024 , 41(1) : 60 -70 . DOI: 10.13866/j.azr.2024.01.06
表1 研究样地植被群落特征(10 m×10 m样方内)Tab. 1 Characteristics of vegetation communities in different micro-topography |
| 样地部位 | 物种 | 株高/cm | 东西长/cm | 南北长/cm | 覆盖度/% | 密度/株 |
|---|---|---|---|---|---|---|
| 沙丘顶部 | 樟子松 | 57.30±16.89Ba | 35.30±7.54Ba | 36.40±10.82Ba | 0.13±0.06Ba | 19 |
| 沙棘 | 82.20±30.20Ba | 81.10±25.44ABa | 84.70±25.91ABa | 0.73±0.40ABa | 63 | |
| 小叶杨 | 195.50±145.13Aa | 120.00±104.73Aa | 118.70±105.31Aa | 2.41±3.51Aa | 11 | |
| 迎风坡中部 | 樟子松 | 50.60±13.88Ba | 26.60±4.45Bb | 30.80±5.94Ba | 0.08±0.02Ba | 10 |
| 沙棘 | 73.40±27.02Ba | 86.70±40.87Aa | 89.10±41.15Aa | 0.91±0.72Aa | 48 | |
| 小叶杨 | 164.60±46.82Aa | 107.30±66.17Aa | 120.50±82.79Aa | 1.78±2.79Aa | 10 | |
| 迎风坡低地 | 樟子松 | 58.50±8.42Ba | 32.30±7.94Bab | 35.50±9.50Ba | 0.11±0.05Ba | 22 |
| 沙棘 | 81.70±22.98Ba | 69.80±21.46Ba | 78.70±13.22ABa | 0.57±0.24Ba | 71 | |
| 小叶杨 | 198.90±105.28Aa | 125.40±77.10Aa | 132.60±84.74Aa | 2.21±2.41Aa | 14 |
注:表中数字表示平均值±标准差,n=10。同列不同大写字母表示相同微地形下不同树种之间差异显著(P<0.05),同列不同小写字母表示同一树种在不同微地形之间差异显著(P<0.05)。 |
| [1] |
冯起, 程国栋. 我国沙地水分分布状况及其意义[J]. 土壤学报, 1999, 36(2): 225-236.
[
|
| [2] |
|
| [3] |
周海, 赵文智, 何志斌. 两种荒漠生境条件下泡泡刺水分来源及其对降水的响应[J]. 应用生态学报, 2017, 28(7): 2083-2092.
[
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
张世才, 王慧娟, 张定海, 等. 腾格里沙漠东南缘3种沙丘4种微地貌上土壤水分与地形-植被因子之间的关系[J]. 甘肃农业大学学报, 2023, 58(3): 160-168, 179.
[
|
| [10] |
朱玉伟, 陈启民, 刘茂秀, 等. 古尔班通古特沙漠南缘沙丘水分的时空分布特征[J]. 草业科学, 2008, 25(12): 6-11.
[
|
| [11] |
田丽慧, 汪海娇, 张登山, 等. 高寒沙地典型固沙植物在沙丘不同地貌部位的水分利用特征[J]. 生态学报, 2021, 41(15): 6215-6226.
[
|
| [12] |
吴汪洋, 张登山, 田丽慧, 等. 青海湖湖东沙地固沙植物的土壤改良作用[J]. 东华理工大学学报(自然科学版), 2022, 45(6): 607-614.
[
|
| [13] |
吴汪洋, 张登山, 田丽慧, 等. 近10年青海湖东沙地人工植被群落特征[J]. 生态学报, 2019, 39(6): 2109-2121.
[
|
| [14] |
汪海娇, 田丽慧, 张登山, 等. 青海湖东沙地不同植被恢复措施下土壤水分变化特征[J]. 干旱区研究, 2021, 38(1): 76-86.
[
|
| [15] |
石明明, 王晓敏, 陈奇, 等. 高寒草地干湿生态系统土壤水分及入渗对降水的响应[J]. 草业学报, 2021, 30(12): 49-58.
[
|
| [16] |
吴汪洋, 张登山, 田丽慧, 等. 青海湖沙地人工治理沙丘的风速廓线变化特征[J]. 水土保持研究, 2013, 20(6): 162-167.
[
|
| [17] |
郑永飞, 陈江峰. 稳定同位素地球化学[M]. 北京: 科学出版社, 2000.
[
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
周海, 郑新军, 唐立松, 等. 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同[J]. 植物生态学报, 2013, 37(7): 665-673.
[
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
黄俊, 吴普特, 赵西宁. 坡面生物调控措施对土壤水分入渗的影响[J]. 农业工程学报, 2010, 26(10): 29-37.
[
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}